作者:黃文功等
來(lái)源:
發(fā)布時(shí)間:2021-10-12
Tag:
點(diǎn)擊:
[麻進(jìn)展] 亞麻響應(yīng)低鉀脅迫轉(zhuǎn)錄譜分析
摘 要:鉀是亞麻生長(zhǎng)發(fā)育必需的大量元素。本研究以鉀高效利用亞麻品種Sofie為試驗(yàn)材料,在低鉀處理12h和96h下,利用轉(zhuǎn)錄組測(cè)序及qRT-PCR進(jìn)行低鉀脅迫下差異基因表達(dá)調(diào)控的研究。結(jié)果表明,低鉀處理7d的亞麻葉片邊緣變黃,與對(duì)照相比,低鉀處理植株矮化。篩選出對(duì)低鉀響應(yīng)強(qiáng)烈的3個(gè)鉀運(yùn)轉(zhuǎn)蛋白基因LusKC1(LusKchannel1)、LusSKOR(LusSTELARK+outwardrectifier)和LusHAK5(LushighaffinityK+transporter5),低鉀脅迫響應(yīng)峰值時(shí)間為12h和96h;與對(duì)照相比,低鉀處理12h鑒定到差異表達(dá)基因1154個(gè)(508個(gè)上調(diào),646個(gè)下調(diào)),GO功能富集分析表明,這些差異表達(dá)基因主要富集于代謝過(guò)程、細(xì)胞進(jìn)程、單一生物過(guò)程、催化活性和結(jié)合功能五大類(lèi),KEGG通路富集分析表明,這些差異表達(dá)基因涉及到能量代謝、碳水化合物代謝、碳代謝、氨基酸代謝、萜類(lèi)化合物代謝和植物激素信號(hào)轉(zhuǎn)導(dǎo)等通路。進(jìn)而篩選出7個(gè)與鉀直接相關(guān)基因(4個(gè)鉀運(yùn)輸?shù)鞍住?個(gè)鉀通道蛋白及1個(gè)鈉鉀鈣交換蛋白)、13個(gè)與激素相關(guān)基因以及6個(gè)與纖維素合成相關(guān)基因。7個(gè)與鉀直接相關(guān)基因中,2個(gè)基因表達(dá)量上調(diào)1.75倍和2.64倍,5個(gè)基因表達(dá)量下調(diào)1.21~9.57倍。以上解析的差異基因初步揭示了亞麻低鉀涉及的轉(zhuǎn)錄調(diào)控途徑,可為亞麻耐低鉀相關(guān)基因的克隆與功能驗(yàn)證奠定基礎(chǔ)。
關(guān)鍵詞:亞麻;低鉀;轉(zhuǎn)錄組;差異表達(dá)基因
中國(guó)耕地土壤鉀含量普遍低,以土壤中速效鉀含量70mgkg-1為標(biāo)準(zhǔn),中國(guó)低鉀土壤總面積約2.27×108hm2,嚴(yán)重制約了作物的正常生長(zhǎng)發(fā)育。中國(guó)鉀肥施用量從2011年的6.05×109kg增加到2017年的1.95×1010kg。大量施用鉀肥,不僅增加生產(chǎn)成本、加速鉀資源枯竭,并造成環(huán)境污染問(wèn)題。研究表明,不同物種甚至同一作物不同品種在土壤鉀素利用上表現(xiàn)出顯著差異。最近,轉(zhuǎn)基因等方法被用來(lái)提高鉀吸收和利用效率。這些措施將緩解鉀素資源的進(jìn)一步枯竭,并提高作物產(chǎn)量和品質(zhì)。亞麻(LinumusttatissimumL.)是一種廣泛種植的經(jīng)濟(jì)作物,有悠久的栽培歷史。鉀是亞麻生長(zhǎng)發(fā)育必需的大量元素。對(duì)亞麻鉀吸收和利用進(jìn)行科學(xué)研究,提高亞麻對(duì)低鉀脅迫適應(yīng)性,對(duì)于中國(guó)農(nóng)業(yè)化肥實(shí)現(xiàn)“雙減”,保護(hù)環(huán)境,避免過(guò)量施用化肥造成的環(huán)境污染,實(shí)現(xiàn)綠水青山理念,實(shí)現(xiàn)中國(guó)生態(tài)農(nóng)業(yè)綠色可持續(xù)發(fā)展有重要意義。
鉀是植物生長(zhǎng)的必需元素,是植物體內(nèi)含量最高的陽(yáng)離子,占植物干物質(zhì)總量2%~10%,在生長(zhǎng)發(fā)育中起重要作用。植物中的鉀調(diào)節(jié)許多物理過(guò)程包括植株生長(zhǎng)、產(chǎn)量和品質(zhì)數(shù)。鉀參與光合作用及葉綠素合成,鉀是植物體中重要的滲透調(diào)節(jié)物質(zhì)。K+遷移對(duì)低鉀土壤中的植物生長(zhǎng)至關(guān)重要,已發(fā)現(xiàn)許多鉀通道,如KT/KUP/HAK轉(zhuǎn)運(yùn)蛋白家族和KCOs家族。還發(fā)現(xiàn)其他具有雙重功能的轉(zhuǎn)運(yùn)體,包括具有Na+和K+轉(zhuǎn)運(yùn)體雙重功能的HKT轉(zhuǎn)運(yùn)體和CBL-CIPK,后者受Ca2+信號(hào)通路調(diào)節(jié),并激活A(yù)KT1通道。此外,ROS信號(hào)通路中的RCI3/RAP被認(rèn)為激活轉(zhuǎn)運(yùn)體HAK。植物低鉀表現(xiàn)明顯癥狀為莖弱、容易倒伏、葉片失水、耐旱性和耐寒性降低、蛋白質(zhì)和葉綠素分解、葉片變黃最終組織壞死。
目前已發(fā)表的亞麻基因組序列圖極大地促進(jìn)了亞麻的研究。亞麻的簡(jiǎn)化基因組測(cè)序結(jié)果表明,遺傳圖譜總長(zhǎng)度為1483.25cM,2339個(gè)標(biāo)記均勻分布于15個(gè)連鎖群,分別對(duì)應(yīng)于亞麻15條染色體,相鄰標(biāo)記間平均距離為0.63cM,該圖譜是目前為止密度最高的亞麻全基因組遺傳連鎖圖譜。有學(xué)者提出一種涉及激活蛋白的低鉀反應(yīng)機(jī)制的存在。目前亞麻對(duì)低鉀脅迫的分子機(jī)制及參與這一調(diào)控過(guò)程的基因仍不明晰。特定響應(yīng)基因表達(dá)的變化決定了鉀脅迫下亞麻形態(tài)及生理變化,差異基因表達(dá)研究有利于揭示鉀脅迫下亞麻內(nèi)在的分子響應(yīng)機(jī)制,為分子輔助品種選育奠定基礎(chǔ)。本研究以亞麻品種Sofie為研究材料,深入挖掘低鉀脅迫下Sofie中的差異表達(dá)基因,通過(guò)揭示鉀脅迫相關(guān)的分子及信號(hào)通路,以期為亞麻耐低鉀相關(guān)基因的克隆與功能驗(yàn)證奠定基礎(chǔ)。
1 材料與方法
1.1 植物材料
鉀高效利用亞麻品種Sofie由國(guó)家麻類(lèi)種質(zhì)改良中心亞麻分中心提供。
1.2 生長(zhǎng)條件及脅迫處理
將Sofie種子播種于裝有蛭石(已經(jīng)過(guò)121℃濕熱滅菌20min)的紙杯中,紙杯置于塑料托盤(pán)上,每個(gè)托盤(pán)澆等體積水,每個(gè)紙杯播種30粒種子。出苗至脅迫處理前每3天澆1/2MS,將播種后的亞麻置于相對(duì)濕度70%的植物培養(yǎng)箱中,23℃,16h光照/18℃,8h黑暗。設(shè)MS培養(yǎng)基中的K+濃度(20mmolL-1K+)和0(0mmolL-1K+)2個(gè)鉀處理梯度。
在Sofie生長(zhǎng)至3對(duì)真葉時(shí)(出苗后7d)進(jìn)行2種濃度的鉀處理,每個(gè)組培瓶中放置10株均勻一致的亞麻植株,每個(gè)組培瓶200mL培養(yǎng)液,處理重復(fù)3次。每3d更換1次培養(yǎng)液,直至出現(xiàn)低鉀癥狀(葉片邊緣變黃)為止。
生長(zhǎng)一致的3周Sofie幼苗正常澆MS培養(yǎng)液(充足的K+供應(yīng),含KCl)作為低鉀對(duì)照(ck),澆低鉀培養(yǎng)液(無(wú)KCl)的作為低鉀處理(lk)。ck營(yíng)養(yǎng)液是改變的MS培養(yǎng)液,KCl代替KNO3。ck和lk脅迫12h和96h,整株苗取樣,液氮凍后80℃貯存,供RNA提取用于后續(xù)測(cè)序分析。
1.3 qRT-PCR引物
根據(jù)擬南芥中的鉀運(yùn)轉(zhuǎn)蛋白相關(guān)基因,在網(wǎng)站(http://phytozome.jgi.doe.gov/pz/portal.htmL)設(shè)計(jì)亞麻L(zhǎng)usKC1、LusSKOR、LusAKT2、LusHAK5、LusKUP2、LusKUP3、LusKUP12、LusKEA5、LusCHX17共9個(gè)基因的引物,每個(gè)基因設(shè)計(jì)3對(duì)引物。
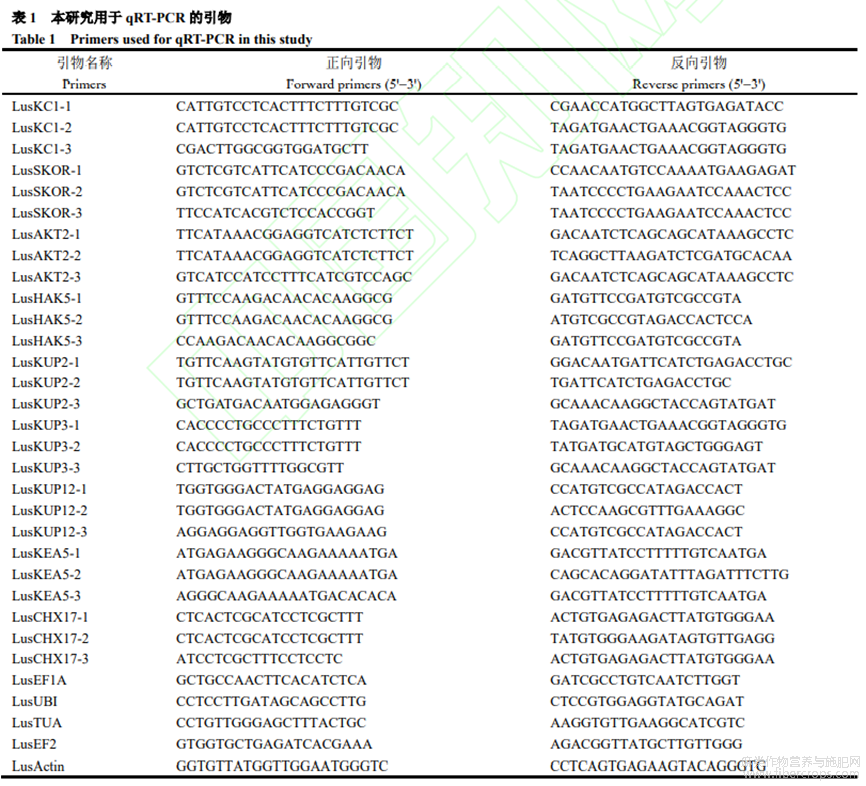
1.4 轉(zhuǎn)錄組測(cè)序
采用TIANGENRNApreppure多糖多酚植物總RNA提取試劑盒提取RNA。用TIANGENTgemSpectrophotometen檢測(cè)所提取RNAA260/A280比值及濃度。ck和lk處理12h和96h提取的Sofie整株植株RNA(3次重復(fù))分別命名為12h-ck-1、12h-ck-2、12h-ck-3、12h-ks-1、12h-ks-2、12h-ks-3、96h-ck-1、96h-ck-2、96h-ck-3、96h-ks-1、96h-ks-2和96h-ks-3。總RNA的質(zhì)量和數(shù)量分析、建庫(kù)及Illumina測(cè)序由華大基因公司(BGI,Shenzhen,China)完成。RNA-seq相應(yīng)數(shù)據(jù)(編號(hào):SRP120129)上傳至公共數(shù)據(jù)庫(kù)NCBI(https://www.ncbi.nlm.nih.gov/)。
1.5 qRT-PCR分析
按上述方法提取總RNA。TIANGENRNApreppurePlantKit已含有去基因組DNA試劑,逆轉(zhuǎn)錄時(shí)省去該步驟。逆轉(zhuǎn)錄方法采用ReverTraAceqPCRRTMasterMixwithgDNARemover(CodeNo.FSQ-301)。RNA的變性:將RNA65℃5min,立即冰上冷卻。逆轉(zhuǎn)錄反應(yīng)體系為10µL,包含RNAtemplate0.5pg~0.5µg、Nuclease-freeWater、5×RTMasterMixⅡ。輕輕攪拌反應(yīng)溶液,以下溫度進(jìn)行反應(yīng):37℃15min,50℃5min,98℃5min,4℃保存。
將1μg總RNA合成第1鏈cDNA。PCR反應(yīng)體系為95℃3min;95℃5s,60℃40s,72℃10s,45個(gè)循環(huán)。為驗(yàn)證引物特異性,45個(gè)循環(huán)后進(jìn)行溶解曲線繪制。qRT-PCR試驗(yàn)設(shè)置3次生物學(xué)重復(fù)。qRT-PCR試驗(yàn)內(nèi)參基因?yàn)長(zhǎng)usActin。利用2−ΔΔCt計(jì)算基因相對(duì)表達(dá)量。利用Primer3(http://frodo.wi.mit.edu/primer3)設(shè)計(jì)引物。
1.6 統(tǒng)計(jì)分析
所有試驗(yàn)3次重復(fù),數(shù)據(jù)用平均值±標(biāo)準(zhǔn)差(SD)表示。采用SPSS17.0軟件分析數(shù)據(jù),各組間比較采用單因素方差分析。P<0.05的值為顯著差異。
2 結(jié)果與分析
2.1 亞麻低鉀脅迫響應(yīng)時(shí)間的確定
2.1.1亞麻低鉀脅迫處理前后亞麻表型
Sofie出苗7d,在蛭石中的生長(zhǎng)狀態(tài)及各植株生長(zhǎng)情況見(jiàn)圖1-A。由圖1-B可看出,各植株生長(zhǎng)一致,長(zhǎng)勢(shì)良好。處理時(shí),對(duì)照和處理的生長(zhǎng)狀態(tài)見(jiàn)圖1-C。處理7d,低鉀和對(duì)照的生長(zhǎng)狀態(tài)見(jiàn)圖1-D和E。說(shuō)明低鉀處理7d植株出現(xiàn)葉片邊緣變黃癥狀,和對(duì)照相比,低鉀處理植株矮化。

2.1.2 亞麻低鉀脅迫標(biāo)記基因篩選
以低鉀處理48hK0的cDNA稀釋10倍為模板進(jìn)行PCR,篩選標(biāo)記基因引物及內(nèi)參。從圖2-A~C可看出,引物L(fēng)usKC1-1、LusSKOR-1、LusHAK5-2、LusKEA5-2和LusActin特異性好。本試驗(yàn)從4個(gè)特異性好的標(biāo)記基因引物L(fēng)usKC1-1、LusSKOR-1、LusHAK5-2和LusKEA5-2中選取3個(gè)標(biāo)記基因LusKC1、LusSKOR、LusHAK5及內(nèi)參LusActin進(jìn)行qRT-PCR試驗(yàn)。

2.1.3亞麻低鉀脅迫響應(yīng)時(shí)間篩選
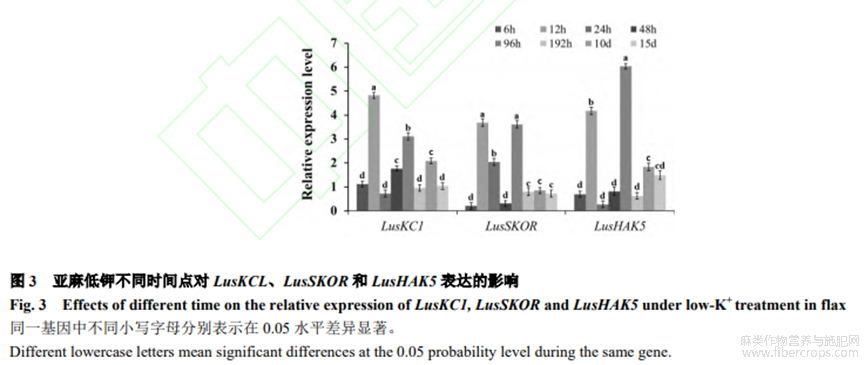
在低鉀脅迫6h、12h、24h、48h、96h、192h、10d和15d時(shí),利用篩出的標(biāo)記基因引物L(fēng)usKC1-1、LusSKOR-1、LusHAK5-2及內(nèi)參LusActin引物進(jìn)行qRT-PCR試驗(yàn)。從圖3可看出,LusKC1在12h和96h表達(dá)量顯著高于其他時(shí)間(P<0.05);LusSKOR在12h和96h表達(dá)量高,這2個(gè)時(shí)間LusSKOR表達(dá)量顯著高于其他時(shí)間(P<0.05);LusHAK5在12h和96h表達(dá)量顯著高于其他時(shí)間(P<0.05)。綜合3個(gè)標(biāo)記基因LusKC1、LusSKOR和LusHAK5qRT-PCR表達(dá)情況發(fā)現(xiàn),3個(gè)標(biāo)記基因12h開(kāi)始高表達(dá),96h表達(dá)處于另一個(gè)高峰。選取低鉀處理及對(duì)照在12h和96h取樣,進(jìn)行后續(xù)轉(zhuǎn)錄組測(cè)序。
2.2 亞麻低鉀的RNA-seq
2.2.1 亞麻低鉀轉(zhuǎn)錄組測(cè)序統(tǒng)計(jì)分析
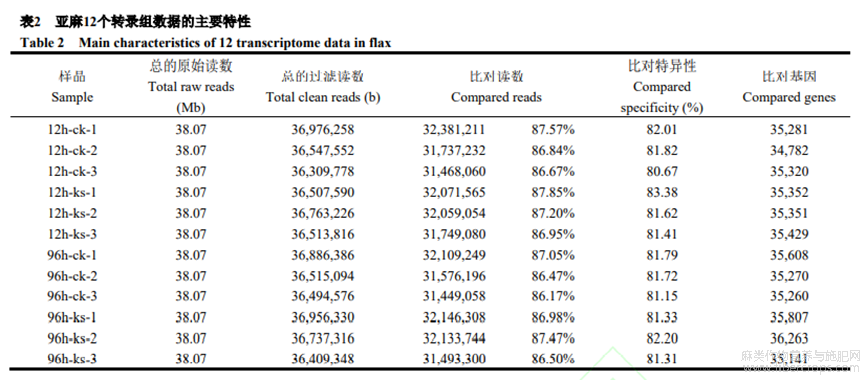
利用低鉀12h和96hSofie及對(duì)應(yīng)時(shí)間對(duì)照樣品總RNA構(gòu)建illumina測(cè)序文庫(kù)。每個(gè)文庫(kù)平均得到38.07Mb原始測(cè)序讀數(shù),其中95.38%以上為過(guò)濾讀數(shù)。將過(guò)濾讀數(shù)與亞麻基因組序列比對(duì),至少86.17%讀數(shù)成功與數(shù)據(jù)庫(kù)參考基因組序列比對(duì)上(表2)。
2.2.2 亞麻低鉀差異表達(dá)基因篩選
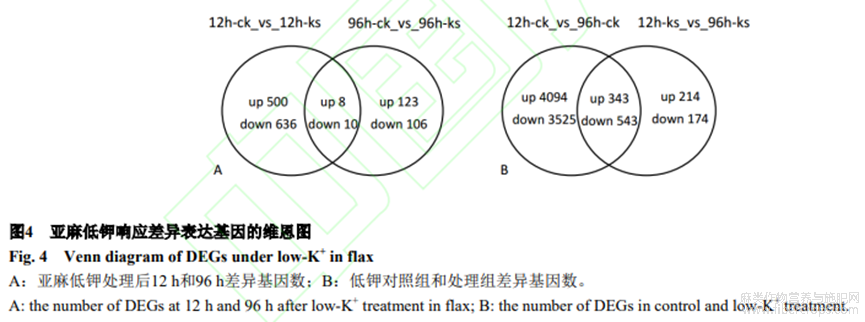
本研究將差異表達(dá)基因的差異倍數(shù)閾值設(shè)為≥2.0,q-value閾值設(shè)為≤0.05。利用q-value閾值<0.05和log2ratio的絕對(duì)值≥1來(lái)篩選出低鉀的差異表達(dá)基因。低鉀12h鑒定到1154個(gè)(508個(gè)上調(diào)表達(dá),646個(gè)下調(diào)表達(dá))低鉀響應(yīng)表達(dá)基因。低鉀96h差異表達(dá)基因明顯減少,共鑒定到247個(gè)(131個(gè)上調(diào)表達(dá),116個(gè)下調(diào)表達(dá))低鉀響應(yīng)表達(dá)基因。圖4-A展示了不同低鉀時(shí)期的差異表達(dá)基因的異同情況,18個(gè)基因在兩時(shí)期均響應(yīng),其中上調(diào)基因8個(gè),下調(diào)基因10個(gè),推測(cè)這些基因可能在整個(gè)低鉀時(shí)期有重要功能。圖4-B顯示低鉀對(duì)照組和處理組差異基因數(shù)。比較這些結(jié)果消除了基因表達(dá)與時(shí)間和生長(zhǎng)相關(guān)的自然變異。在ck組和ks組中,有886個(gè)基因共同表達(dá),其中343個(gè)基因上調(diào),543個(gè)基因下調(diào)。
2.3 GO功能富集
對(duì)差異表達(dá)基因(1154個(gè))進(jìn)行GO功能富集發(fā)現(xiàn),在12h-ksvs12h-ck對(duì)比組差異基因主要富集于代謝過(guò)程、細(xì)胞進(jìn)程、單一生物過(guò)程、催化活性和結(jié)合功能五大類(lèi)(圖5)。
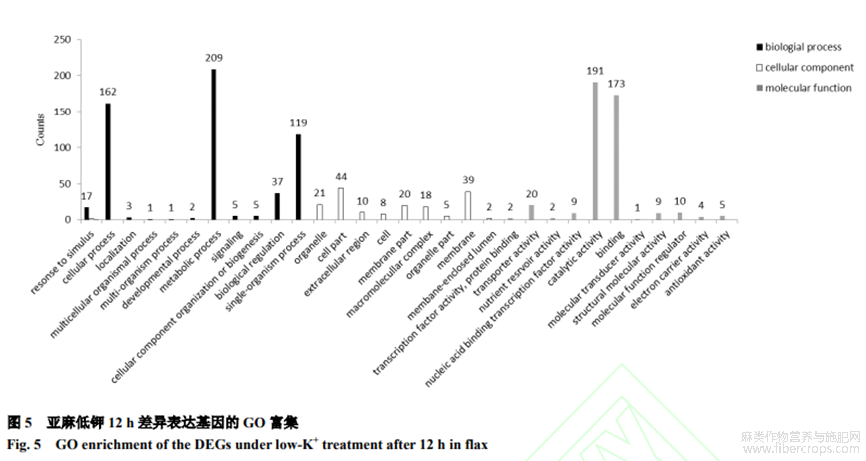
2.4 KEGG通路富集
為進(jìn)一步理解差異表達(dá)基因的功能,將12h-ksvs12h-ck對(duì)比組所有差異基因(1154個(gè))與KEGG數(shù)據(jù)庫(kù)比對(duì),確定差異表達(dá)基因參與的最主要的能量代謝(光合有機(jī)物固碳、光合作用),碳水化合物代謝(戊糖、果糖磷酸途徑、甘露糖代謝)、碳代謝、氨基酸代謝、萜類(lèi)化合物代謝和植物激素信號(hào)轉(zhuǎn)導(dǎo)通路(圖6)。差異表達(dá)基因中有本研究關(guān)注的pathway注釋的基因?yàn)?個(gè),富集的途徑有5條,包括光合作用途徑(photosynthesis)1條、植物激素信號(hào)轉(zhuǎn)導(dǎo)途徑(planthormonesignaltransduction)3條及萜類(lèi)主鏈生物合成途徑(Terpenoidsbackbonebiosynthesis)1條。其中基因Potassiumtransporter5-like(MSTRG.20565.1)表達(dá)量上調(diào)2.64倍,在光合作用途徑中負(fù)責(zé)K+運(yùn)輸;基因TIR(MSTRG.7397.8)表達(dá)量上調(diào)8.52倍,在生長(zhǎng)素合成途徑中負(fù)責(zé)生長(zhǎng)素的合成;基因AHP1(MSTRG.25736.1)表達(dá)量下調(diào)1.81倍,在細(xì)胞分裂素合成途徑中負(fù)責(zé)細(xì)胞分裂素的合成;基因CTR1(MSTRG.16160.5)表達(dá)量下調(diào)6.23倍,在乙烯合成途徑中負(fù)責(zé)乙烯的合成;基因FEI1(MSTRG.5636.3)表達(dá)量下調(diào)1.86倍,在萜類(lèi)主鏈生物合成途徑中負(fù)責(zé)細(xì)胞壁的合成。
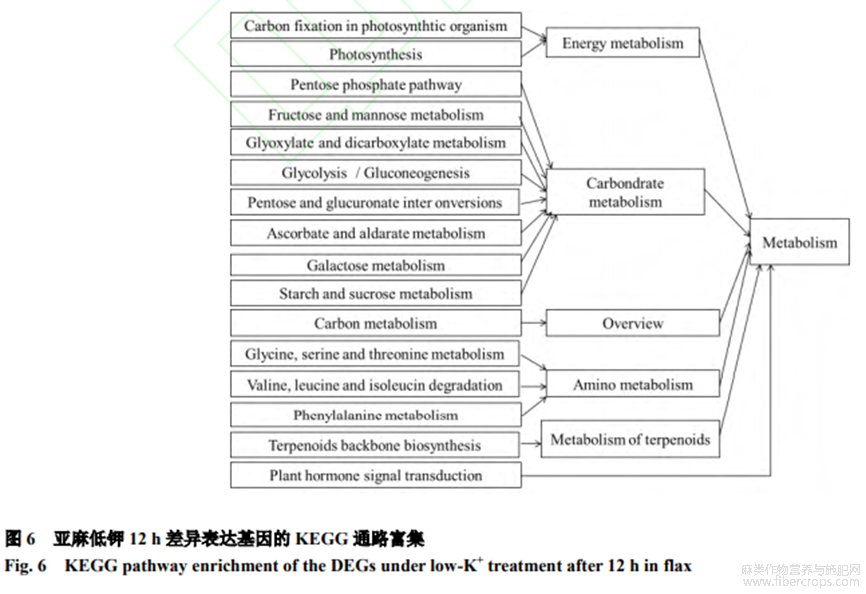
2.5 差異表達(dá)基因的功能分析
2.5.1 亞麻低鉀下與鉀直接相關(guān)基因篩選
本研究在亞麻低鉀轉(zhuǎn)錄組數(shù)據(jù)的差異基因中篩選出7個(gè)與鉀直接相關(guān)基因(表3),包括4個(gè)鉀運(yùn)輸?shù)鞍?MSTRG.20565.1、MSTRG.24915.2、MSTRG.10817.1和MSTRG.6817.1),2個(gè)鉀通道蛋白(MSTRG.4695.1和MSTRG.30540.1)及1個(gè)鈉鉀鈣交換蛋白(MSTRG.14498.2)。其中2個(gè)上調(diào)基因(MSTRG.20565.1和MSTRG.4695.1),5個(gè)下調(diào)基因(MSTRG.24915.2、MSTRG.10817.1、MSTRG.30540.1、MSTRG.14498.2和MSTRG.6817.1),這些基因可能在鉀脅迫響應(yīng)中起重要作用。
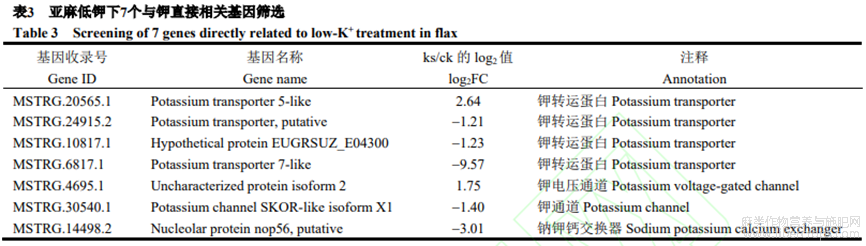
本研究發(fā)現(xiàn),許多差異表達(dá)基因在低鉀脅迫信號(hào)轉(zhuǎn)導(dǎo)和離子轉(zhuǎn)運(yùn)體調(diào)控通路中扮演重要角色,如附圖1。1個(gè)AKT通道蛋白(上調(diào)2.15倍),參與鉀通道轉(zhuǎn)運(yùn),響應(yīng)于非生物脅迫;19個(gè)CIPK信號(hào)轉(zhuǎn)導(dǎo)蛋白(上調(diào)1.46~11.76倍),參與鈣信號(hào)傳導(dǎo),響應(yīng)于非生物脅迫;1個(gè)CML信號(hào)轉(zhuǎn)導(dǎo)蛋白(上調(diào)3.74倍),參與鈣信號(hào)傳導(dǎo),影響植物生理代謝;2個(gè)ARF信號(hào)轉(zhuǎn)導(dǎo)蛋白(下調(diào)3.25倍和4.61倍),參與生長(zhǎng)素調(diào)控,影響側(cè)根發(fā)育;5個(gè)NRT轉(zhuǎn)運(yùn)蛋白(下調(diào)1.14~4.69倍),參與硝酸鹽轉(zhuǎn)運(yùn),影響植物的生長(zhǎng)發(fā)育。鑒于這些基因直接參與鉀信號(hào)傳導(dǎo)及K+運(yùn)輸,與Wang等報(bào)道一致。推測(cè)低鉀脅迫將直接影響這些基因的表達(dá),進(jìn)而影響亞麻中鉀信號(hào)的傳導(dǎo)及K+運(yùn)輸,從而影響亞麻的表型。
2.5.2 亞麻低鉀下與激素相關(guān)基因篩選
在所有差異對(duì)比組中,共103個(gè)基因注釋到激素響應(yīng)蛋白功能,包括17個(gè)ABA響應(yīng)蛋白、16個(gè)乙烯響應(yīng)蛋白、22個(gè)生長(zhǎng)素響應(yīng)蛋白、10個(gè)JA響應(yīng)蛋白以及38個(gè)細(xì)胞分裂素響應(yīng)蛋白。本研究發(fā)現(xiàn),13個(gè)與激素直接相關(guān)的基因(表4),其中1個(gè)TIR(上調(diào)8.52倍),一種植物生長(zhǎng)素受體,參與植物生長(zhǎng)調(diào)控;2個(gè)AUX(下調(diào)1.44倍和1.51倍),生長(zhǎng)素響應(yīng)基因,應(yīng)答生長(zhǎng)素效應(yīng);1個(gè)ARF(下調(diào)9.62倍),生長(zhǎng)素初期響應(yīng)基因;2個(gè)GH3(下調(diào)3.02倍和3.58倍),植物生長(zhǎng)素原初反應(yīng)基因,參與植物生長(zhǎng)素調(diào)節(jié);1個(gè)AHP(下調(diào)1.81倍),一種組氨酸磷酸轉(zhuǎn)移蛋白,參與細(xì)胞分裂素的負(fù)向調(diào)控;1個(gè)EIN2(下調(diào)10.02倍),乙烯信號(hào)關(guān)鍵基因,參與乙烯信號(hào)通路的正向調(diào)控;2個(gè)EBF(下調(diào)1.20倍和3.10倍),負(fù)反饋調(diào)控EIN3,參與乙烯信號(hào)轉(zhuǎn)導(dǎo);3個(gè)CTR1(下調(diào)6.23倍、1.90倍和7.72倍),介導(dǎo)乙烯受體的信號(hào)負(fù)調(diào)控乙烯反應(yīng)。
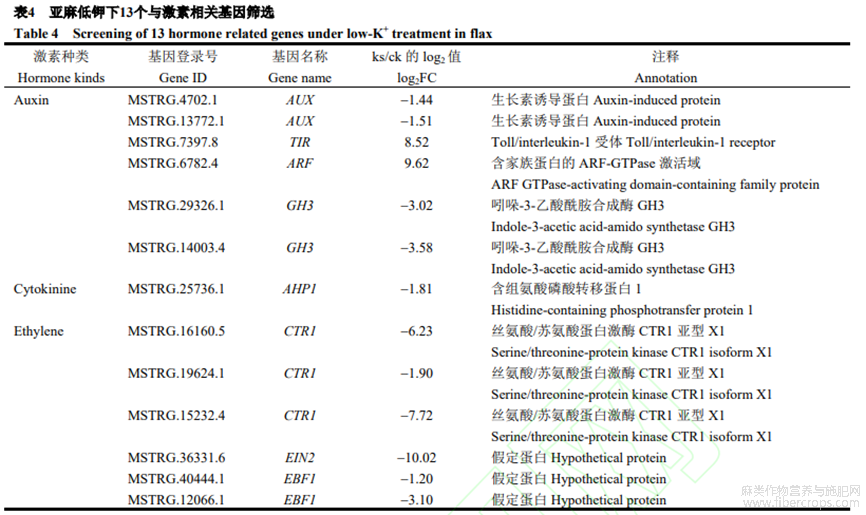
本研究篩選出生長(zhǎng)素合成通路6個(gè)基因,包括1個(gè)TIR、2個(gè)AUX、1個(gè)ARF、2個(gè)GH3,低鉀脅迫下,這些基因調(diào)控亞麻植株生長(zhǎng),造成植株矮化;細(xì)胞分裂素合成通路1個(gè)基因AHP,低鉀脅迫下,AHP調(diào)控細(xì)胞分裂,造成亞麻植株矮化的表型;乙烯合成通路6個(gè)基因,包括3個(gè)CTR、1個(gè)EIN、2個(gè)EBF,低鉀脅迫下,這些基因引起亞麻植株衰老,即葉片變黃(附圖2)。這3個(gè)通路為揭示植物激素對(duì)低鉀脅迫反應(yīng)分子機(jī)制奠定基礎(chǔ)。低鉀脅迫可能誘導(dǎo)這些基因差異表達(dá),調(diào)控激素合成,進(jìn)而影響亞麻的生長(zhǎng)發(fā)育。
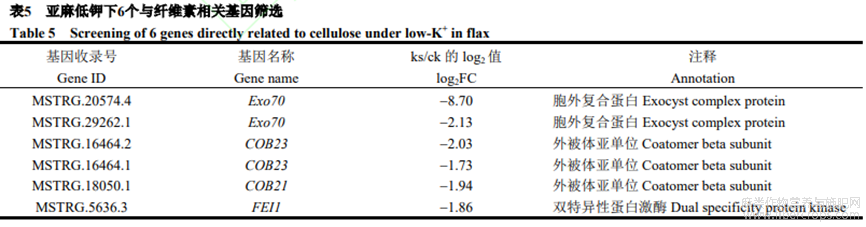
2.5.3 亞麻低鉀下與纖維素相關(guān)基因篩選
本研究在差異表達(dá)基因中篩選出6個(gè)與纖維素相關(guān)基因(表5),包括2個(gè)Exo70(下調(diào)8.70倍和2.13倍),組成胞泌復(fù)合體的關(guān)鍵亞基,參與復(fù)合體在靶膜組裝;2個(gè)COB23(下調(diào)2.03倍和1.73倍)及1個(gè)COB21(下調(diào)1.94倍),植物糖磷脂酰肌醇錨定蛋白,定向微纖絲,突變后阻礙纖維素合成;1個(gè)FEI1(下調(diào)1.86倍),一種雙特異性蛋白激酶,參與纖維素合成。本研究篩選出2個(gè)Exo70、2個(gè)COB23、1個(gè)COB21和1個(gè)FEI1(附圖3),鑒于這些基因直接參與纖維素合成,低鉀脅迫可能誘導(dǎo)這些基因表達(dá),抑制亞麻纖維素合成,進(jìn)而調(diào)控產(chǎn)量形成。
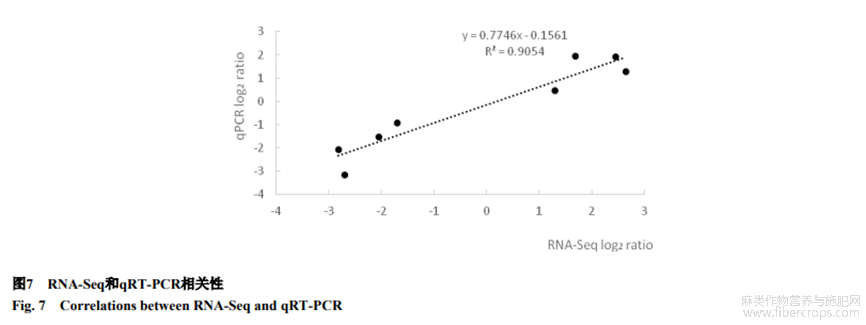
2.6 RNA-Seq和qRT-PCR相關(guān)性
為驗(yàn)證轉(zhuǎn)錄組測(cè)序分析結(jié)果,隨機(jī)選取Lus10024152、Lus10021422、Lus10028689、Lus10040830、Lus10024511、Lus10013304、Lus10024074和Lus100090928個(gè)基因用于qRT-PCR分析。2種方法得到的基因表達(dá)變化數(shù)據(jù)相關(guān)系數(shù)R2達(dá)到0.91(圖7),說(shuō)明RNA-seq測(cè)序數(shù)據(jù)相對(duì)準(zhǔn)確,低鉀相關(guān)基因表達(dá)量結(jié)果可信。
3 討論
3.1 低鉀對(duì)亞麻表型的影響及亞麻響應(yīng)低鉀的時(shí)間
植物長(zhǎng)期受到低鉀脅迫時(shí),會(huì)出現(xiàn)黃化病,老葉先顯現(xiàn)癥狀。植物低鉀影響莖葉及根的生長(zhǎng)發(fā)育。植株莖葉低鉀早期老葉葉尖失綠。隨低鉀時(shí)間增加,萎黃癥狀從老葉轉(zhuǎn)移到新葉。嚴(yán)重低鉀時(shí),植物葉子從葉尖到整個(gè)葉緣顯現(xiàn)出萎黃病,最終壞死。隨腐胺在葉片中積累,植物葉片萎黃并發(fā)生大量褐斑。與上述植物低鉀表型相似,低鉀亞麻植株出現(xiàn)葉片邊緣變黃癥狀,并表現(xiàn)出矮化的表型。
研究表明,西瓜低鉀響應(yīng)早期和后期的時(shí)間點(diǎn)分別為6h和120h。而本研究通過(guò)亞麻低鉀8個(gè)時(shí)間點(diǎn)篩選出12h和96h為亞麻低鉀響應(yīng)的早期和后期時(shí)間點(diǎn)。與西瓜低鉀響應(yīng)不同的是,亞麻低鉀響應(yīng)早期時(shí)間(12h)晚于西瓜(6h),但亞麻后期響應(yīng)時(shí)間(96h)早于西瓜(120h)。
3.2 植物低鉀脅迫的信號(hào)轉(zhuǎn)導(dǎo)和離子轉(zhuǎn)運(yùn)體調(diào)控
本研究2個(gè)ARF信號(hào)轉(zhuǎn)導(dǎo)蛋白(下調(diào)3.25倍和4.61倍),參與生長(zhǎng)素調(diào)控,影響側(cè)根發(fā)育;5個(gè)NRT轉(zhuǎn)運(yùn)蛋白(下調(diào)1.14~4.69倍),參與硝酸鹽轉(zhuǎn)運(yùn),影響植物的生長(zhǎng)發(fā)育。鑒于這些基因直接參與鉀信號(hào)傳導(dǎo)及鉀離子運(yùn)輸,與Wang等報(bào)道一致。推測(cè)低鉀脅迫將直接影響這些基因的表達(dá),進(jìn)而影響亞麻中鉀信號(hào)的傳導(dǎo)及K+運(yùn)輸,從而影響亞麻的表型。
鉀是植物生長(zhǎng)所需的主要營(yíng)養(yǎng)物質(zhì)。對(duì)低K+條件的適應(yīng)性機(jī)制主要為鈣調(diào)磷酸酶B樣蛋白(CalcineurinB-likeCalciumSensors,CBL)和CBL相互作用激酶(CBL-InteractingProteinKinases,CIPK)組成的Ca2+信號(hào)網(wǎng)絡(luò)。低鉀脅迫信號(hào)轉(zhuǎn)導(dǎo)和離子轉(zhuǎn)運(yùn)體調(diào)控通路涉及大量膜上K+信號(hào)轉(zhuǎn)導(dǎo)蛋白和離子通道基因及細(xì)胞質(zhì)和核中一些信號(hào)轉(zhuǎn)導(dǎo)蛋白及離子轉(zhuǎn)運(yùn)蛋白。低鉀脅迫通過(guò)調(diào)控細(xì)胞質(zhì)和細(xì)胞核中一些信號(hào)轉(zhuǎn)導(dǎo)蛋白影響膜上相關(guān)信號(hào)轉(zhuǎn)導(dǎo)蛋白和離子通道基因表達(dá)(附圖1)。本研究低鉀脅迫發(fā)現(xiàn)大量差異表達(dá)基因,AKT通道蛋白在亞麻低鉀處理后其表達(dá)量上調(diào)2.15倍,而擬南芥中CIPK23及KC1協(xié)同調(diào)控AKT1介導(dǎo)低鉀脅迫,水稻中過(guò)表達(dá)AKT1可通過(guò)提高組織內(nèi)K+含量提高植株對(duì)滲透和干旱脅迫抗性,說(shuō)明AKT通道蛋白在亞麻和擬南芥或水稻有類(lèi)似功能;發(fā)現(xiàn)CIPK信號(hào)轉(zhuǎn)導(dǎo)蛋白在亞麻低鉀處理后其表達(dá)量上調(diào)1.46~11.76倍,而楊樹(shù)中CBL-CIPK信號(hào)通路可通過(guò)調(diào)控Shaker-like鉀通道增加對(duì)低鉀脅迫耐受性,說(shuō)明CIPK信號(hào)通路在亞麻和楊樹(shù)有類(lèi)似功能;發(fā)現(xiàn)ARF信號(hào)轉(zhuǎn)導(dǎo)蛋白在亞麻低鉀處理后其表達(dá)量下調(diào)3.25倍和4.61倍,而擬南芥中低鉀脅迫下ARF2磷酸化可解除對(duì)K+轉(zhuǎn)運(yùn)基因HAK5抑制,促進(jìn)鉀吸收,說(shuō)明ARF信號(hào)轉(zhuǎn)導(dǎo)蛋白在亞麻和擬南芥有類(lèi)似功能;發(fā)現(xiàn)NRT轉(zhuǎn)運(yùn)蛋白在亞麻低鉀處理后其表達(dá)量下調(diào)1.14~4.69倍,而擬南芥中NPF7.3/NRT1.5介導(dǎo)的NO3-遷移可通過(guò)調(diào)控SKOR影響K+從根到芽遷移,說(shuō)明NRT轉(zhuǎn)運(yùn)蛋白在亞麻和擬南芥有類(lèi)似功能。鑒于這些基因在低鉀脅迫亞麻中的表達(dá)明顯改變,推測(cè)這些基因在亞麻對(duì)低鉀脅迫的響應(yīng)中發(fā)揮著重要作用。
3.3 植物激素在低鉀脅迫中的作用
植物激素廣泛參與逆境生理和生化反應(yīng)。乙烯是植物對(duì)非生物脅迫應(yīng)答中重要信號(hào)分子之一,它與乙烯受體等主要轉(zhuǎn)錄因子結(jié)合,激活下游基因表達(dá),觸發(fā)乙烯應(yīng)答。生長(zhǎng)素可誘導(dǎo)ARF/AUX/GH3等基因在植物脅迫應(yīng)答中快速瞬時(shí)高表達(dá)。脫落酸作為植物地下-地上部分之間信息傳遞的中心媒介,通過(guò)激活K+進(jìn)出細(xì)胞路徑誘導(dǎo)細(xì)胞膨脹壓變化。植物激素也相互影響,乙烯和生長(zhǎng)素通過(guò)抑制主根生長(zhǎng)而促進(jìn)根毛伸長(zhǎng),從而對(duì)低鉀脅迫下根系形態(tài)發(fā)育起指導(dǎo)作用。生長(zhǎng)在低鉀條件的植物根形態(tài)類(lèi)似于生長(zhǎng)在響應(yīng)外源乙烯和生長(zhǎng)素處理的形態(tài)。低鉀條件下,乙烯合成和信號(hào)轉(zhuǎn)導(dǎo)相關(guān)基因表達(dá)增強(qiáng)。
4 結(jié)論
利用qRT-PCR篩選出亞麻對(duì)低鉀響應(yīng)強(qiáng)烈的3個(gè)標(biāo)記基因及內(nèi)參分別為:LusKC1、LusSKOR、LusHAK5和LusActin。鉀脅迫相關(guān)基因?qū)Φ外涰憫?yīng)高峰時(shí)間出現(xiàn)在12h和96h。通過(guò)低鉀轉(zhuǎn)錄組數(shù)據(jù)的分析和挖掘,確定7個(gè)與鉀直接相關(guān)基因(4個(gè)鉀運(yùn)輸?shù)鞍住?個(gè)鉀通道蛋白及1個(gè)鈉鉀鈣交換蛋白),低鉀脅迫直接影響亞麻這些基因的表達(dá),進(jìn)而影響亞麻中鉀信號(hào)的傳導(dǎo)及K+運(yùn)輸;確定13個(gè)與激素相關(guān)基因(生長(zhǎng)素合成通路6個(gè)基因、細(xì)胞分裂素合成通路1個(gè)基因和乙烯合成通路6個(gè)基因),低鉀脅迫誘導(dǎo)這些基因表達(dá),調(diào)控激素合成,進(jìn)而影響亞麻生長(zhǎng)發(fā)育;確定6個(gè)與纖維素合成相關(guān)基因(2個(gè)EXOCYST、3個(gè)COB和1個(gè)FEI基因),低鉀脅迫誘導(dǎo)這些基因表達(dá)抑制亞麻纖維素合成,進(jìn)而調(diào)控產(chǎn)量形成。
文章來(lái)源:黃文功,姜衛(wèi)東,姚玉波,宋喜霞,劉巖,陳思,趙東升,吳廣文,袁紅梅,任傳英,孫中義,吳建忠,康慶華.亞麻響應(yīng)低鉀脅迫轉(zhuǎn)錄譜分析[J/OL].作物學(xué)報(bào):1-13[2021-02-17].http://kns.cnki.net/kcms/detail/11.1809.S.20201215.0849.006.html.
更多閱讀
文章排行
最新文章
Copyright by Ramie Research Institute of Hunan Agricultural University
湖南農(nóng)業(yè)大學(xué)農(nóng)學(xué)院/苧麻研究所 版權(quán)所有 2009-2016 湘ICP備10006838號(hào)-1
