作者:陳文濤等
來源:
發(fā)布時間:2021-10-13
Tag:
點擊:
[麻進展]紅麻HcMS1基因的克隆、表達與功能分析
摘 要:為挖掘紅麻雄性不育核基因,解釋紅麻花蕾敗育的分子機理,通過分析轉(zhuǎn)錄組數(shù)據(jù),利用同源克隆的方法從紅麻花藥中克隆出擬南芥MALE STERILITY1(MS1)的同源不育基因,并命名為HcMS1基因。利用生物信息學方法對HcMS1蛋白進行結(jié)構(gòu)、理化性質(zhì)及親緣關(guān)系等分析,并通過qRT-PCR分析HcMS1基因在紅麻不育系、保持系不同時期的表達水平;構(gòu)建過表達載體,利用葉盤法轉(zhuǎn)化本氏煙草并對轉(zhuǎn)基因煙草進行表型觀察。HcMS1基因序列開放閱讀框(ORF)為1 950bp,編碼649個氨基酸。生物信息學分析顯示,HcMS1蛋白為親水蛋白,定位在細胞核上且含有典型PHD-finger結(jié)構(gòu)域,沒有信號肽和跨膜區(qū)結(jié)構(gòu),與陸地棉MS1蛋白親緣關(guān)系最近,其次為木槿MS1蛋白。qRT-PCR結(jié)果表明,HcMS1基因的表達量在保持系和不育系中均呈現(xiàn)先上升、后下降的趨勢,且在紅麻花蕾的四分體至單核期表達量最高,這與擬南芥中MS1基因表達模式一致;另外在保持系中基因的表達水平顯著高于不育系,推測紅麻敗育與HcMS1基因的低量表達相關(guān)。通過遺傳轉(zhuǎn)化試驗發(fā)現(xiàn),HcMS1轉(zhuǎn)基因煙草株型較矮,花萼大小不一,花筒長度縮短,并出現(xiàn)自交不結(jié)實的現(xiàn)象,說明紅麻HcMS1基因的異源表達有影響本氏煙草正常育性的功能。從紅麻花藥中成功克隆出核不育相關(guān)基因HcMS1,并對其進行功能分析,為后續(xù)紅麻雄性不育育種工作提供了一定的理論依據(jù)與基礎(chǔ)。
關(guān)鍵詞:紅麻雄性不育;HcMS1基因;生物信息學分析;表達分析;煙草轉(zhuǎn)化
紅麻(Hibiscus cannabinus L.)是錦葵科木槿屬一年生纖維作物,具有抗旱、耐鹽堿、適應性廣、生物量大等特性,在造紙、紡織、建筑材料、復合材料、水土保持、吸附劑、飼料、栽培基質(zhì)等方面有良好的應用價值,還可有效地緩解土壤重金屬污染問題。
紅麻有明顯的雜種優(yōu)勢,雜交紅麻的韌皮纖維或生物產(chǎn)量可提高40%左右。雄性不育是利用雜種優(yōu)勢的一條有效途徑,利用細胞質(zhì)雄性不育(Cytoplas-mic male sterility)構(gòu)建三系配套進行雜交制種可以免去人工去雄,同時提高雜交種子的純度。目前,國內(nèi)關(guān)于紅麻雄性不育分子機理的研究工作已陸續(xù)開展。趙艷紅構(gòu)建紅麻線粒體atp9基因過表達載體轉(zhuǎn)化煙草,發(fā)現(xiàn)在陽性植株出現(xiàn)葉形、株高、花冠性狀異常且花藥不正常的現(xiàn)象,另外對線粒體基因atp9、atp6和atpA的RNA編輯研究發(fā)現(xiàn)轉(zhuǎn)變的氨基酸大多以非極性氨基酸取代極性氨基酸。史奇奇以紅麻不育系UG93A和保持系UG93B花藥為樣品,分別建立smallRNA文庫,并使用高通量測序技術(shù)鑒定42個保守miRNA和41個新的miRNA。韋范通過乙酰化蛋白質(zhì)組學分析在UG93A和UG93B花藥中鑒定到56個乙?;揎棽町惖鞍祝l(fā)現(xiàn)乙?;蛘{(diào)控GAPDH酶活力以及組蛋白乙?;{(diào)控了細胞質(zhì)雄性不育的發(fā)生。李增強等通過研究紅麻不育系UG93A和保持系UG93B葉片和不同時期花藥基因組DNA甲基化情況,發(fā)現(xiàn)甲基化變化的基因在不育系和保持系之間的表達差異顯著,推測這些甲基化變化在雄性不育中發(fā)揮了重要的作用。錢景華、陳勵虹等分別克隆紅麻轉(zhuǎn)錄因子MYB21基因和編碼生長素受體的TIR1基因,分析基因表達模式并構(gòu)建了過表達載體和沉默載體。Chen等通過對紅麻不育系、保持系的轉(zhuǎn)錄組測序檢測到838個顯著上調(diào)基因和528個顯著下調(diào)基因,并將其注釋到155個GO和74條KEGG代謝通路上。隨著對紅麻不育機理的研究增多,更多的不育基因得到挖掘,復雜的作用機制和調(diào)控網(wǎng)絡(luò)逐漸清晰,將在紅麻生產(chǎn)實踐發(fā)揮重要的作用。
絨氈層參與花粉壁物質(zhì)合成,為小孢子發(fā)育提供營養(yǎng)并適時地分泌胼胝質(zhì)酶以釋放小孢子,其在花藥發(fā)育中發(fā)揮著重要的作用。DYT1-TDF1-AMS-MS188-MS1遺傳途徑被證實調(diào)控擬南芥絨氈層發(fā)育,其中DYT1、TDF1和AMS在絨氈層早期發(fā)育中起作用,而MS188和MS1在絨氈層晚期發(fā)育中起重要作用。MS1是MS188的直接靶點,在調(diào)控擬南芥孢子體花粉外殼蛋白基因的表達起關(guān)鍵作用。MS1在擬南芥中編碼 PHD-finger蛋白,在四分體期的絨氈層中表達,其突變體不能產(chǎn)生有活力的花粉,花粉在四分體釋放小孢子后不久退化,絨氈層也出現(xiàn)異??张莼?。本試驗在前期紅麻不育系LC0301A和保持系LC0301B的花藥轉(zhuǎn)錄本中,發(fā)現(xiàn)一個顯著差異表達的基因,其編碼蛋白序列與澳洲棉MS1蛋白有90%一致性、與擬南芥MS1蛋白有50%一致性,命名其為HcMS1基因。通過同源克隆的方法獲得了紅麻HcMS1基因,利用生物信息學預測該基因編碼蛋白的結(jié)構(gòu)、保守域和親緣進化關(guān)系等,通過qRT-PCR對紅麻不育系、保持系不同時期的花藥進行表達量分析,并構(gòu)建過表達載體對本氏煙草進行遺傳轉(zhuǎn)化試驗,以驗證HcMS1基因可能在紅麻不育中發(fā)揮一定的作用。
1材料和方法
1.1試驗材料
1.1.1植物材料
試驗所用紅麻品種不育系LC0301A和保持系LC0301B由中國農(nóng)業(yè)科學院麻類所一年生創(chuàng)新育種團隊提供,種植于麻類所長沙創(chuàng)新試驗基地。根據(jù)李建軍等劃分紅麻花粉發(fā)育時期的標準,按照花蕾的大小分為4個時期:造孢細胞和小孢子母細胞至減數(shù)分裂時期為花蕾一期,四分體到單核期為花蕾二期,雙核期為花蕾三期,花粉成熟期為花蕾四期;取各時期花藥于-80℃冰箱保存。
1.1.2試劑、載體與菌株
TRIzol液,DNAaseI,反轉(zhuǎn)錄試劑盒,TaKaRa Ex Taq® DNA Polymerase,DNA回收試劑盒,pBLUE-T載體、Pcambia2301-ky過表達載體,T4DNA連接酶,各種相關(guān)限制性內(nèi)切酶購于武漢金開瑞生物工程有限公司。SteadyPure植物RNA提取試劑盒,Evo M-MLV反轉(zhuǎn)錄試劑,SYBR Green qPCR試劑盒購于湖南艾科瑞生物工程有限公司。大腸桿菌感受態(tài)細胞DH5α,農(nóng)桿菌菌株GV3101購于長沙天楚生物科技有限公司。
1.2試驗方法
1.2.1總RNA提取及cDNA合成
以不育系LC0301A和保持系LC0301B的花藥為材料,采用T-Izol法提取紅麻總RNA,利用瓊脂糖凝膠電泳檢測RNA完整性;采用TOYOBO公司的逆轉(zhuǎn)錄試劑盒(Re-verTra Ace qPCR RT Kit)進行反轉(zhuǎn)錄合成cDNA以用于基因克隆。使用SteadyPure植物RNA提取試劑盒提取4個時期的不育系、保持系紅麻花藥RNA,使用EvoM-MLV反轉(zhuǎn)錄試劑合成cDNA用于qRT-PCR。
1.2.2紅麻HcMS1基因的PCR擴增及克隆
根據(jù)RNA-seq數(shù)據(jù),分析鑒定紅麻HcMS1基因轉(zhuǎn)錄本,利用NCBI(https://www.ncbi.nlm.nih.gov/orffind-er/)的Open Reading Frame Finder找出紅麻HcMS1的開放閱讀框。用此序列在Primer6.0軟件中設(shè)計特異性克隆引物(表1),以紅麻花蕾cDNA為模板,使用Taq酶擴增HcMS1基因的CDS序列,擴增程序:94℃預變性3min;94℃30s,55℃30s,72℃2min,30個循環(huán);72℃延伸5min。PCR產(chǎn)物經(jīng)1%瓊脂糖凝膠電泳后回收目的條帶連接到pBLUE-T載體,轉(zhuǎn)化到大腸桿菌感受態(tài)DH5α中,37℃,100mg/L氨芐青霉素LB培養(yǎng)基上抗性篩選,挑取陽性克隆進行測序驗證。

1.2.3生物信息學分析
Prot Param(https://web.expasy.org/protparam/)分析蛋白質(zhì)理化性質(zhì)ProtScale(https://web.expasy.org/protscal;www.cbs.dtu.dk/services/TMHMM/)分析蛋白質(zhì)的跨膜結(jié)構(gòu)域;SignalP-5.0Server(http://www.cbs.dtu.dk/services/SignalP/)分析蛋白質(zhì)是否含有信號肽;ProtComp9.0進行亞細胞定位進行分析;Inter-Pro(http://www.ebi.ac.uk/interpro/)分析蛋白質(zhì)結(jié)構(gòu)域;SOPMA分析和預測蛋白質(zhì)的二級結(jié)構(gòu);Phyre2分析預測蛋白質(zhì)的三級結(jié)構(gòu);MEGA7.0軟件構(gòu)建分子進化樹。
1.2.4實時熒光定量
PCR根據(jù)HcMS1序列設(shè)計熒光定量引物(表1),以紅麻GAPDH基因為內(nèi)參基因(表1),以不同樣品反轉(zhuǎn)錄cDNA為模板。反應體系為20μL,包含1μL模板,10μL2×SYBRPremix、10μmol/L上游引物和下游引物各0.4μL、滅菌水補足20μL。定量在BioRadCFX96qPCR儀進行,程序設(shè)置為:95℃預變性2min;95℃5s,60℃30s,40個循環(huán)后進行熔解曲線分析,以確定引物的特異性?;蛳鄬Ρ磉_量采用公式2-ΔΔCt計算,使用SPSS21進行單因素方差分析,利用Origin9進行繪圖。
1.2.5HcMS1基因過表達載體構(gòu)建和遺傳轉(zhuǎn)化
根據(jù)HcMS1基因的ORF序列信息,選用SacⅠ/BamHⅠ內(nèi)切酶為酶切位點并設(shè)計擴增引物,引物序列為MS1-F2/R2(表1),以TA克隆為模板進行HcMS1基因的PCR擴增克隆。SacⅠ/BamHⅠ雙酶切pcam-bia2301-ky載體和HcMS1目的基因,回收HcMS1片段和pcambia2301-ky載體,進行連接反應。反應產(chǎn)物轉(zhuǎn)化大腸桿菌并在50mg/L卡那霉素(Kana)和50mg/L利福平(Rif)抗性平板上進行篩選,挑取單克隆菌落過夜搖菌并進行菌落PCR驗證,將PCR陽性菌落提取質(zhì)粒后進行測序,測序引物為35S-F/2301-F(表1)。將測序后比對成功的過表達載體質(zhì)粒,采用凍融法轉(zhuǎn)入GV3101農(nóng)桿菌并在50mg/L卡那霉素和50mg/L利福平抗性平板上28℃進行篩選,挑取單克隆菌落180r/min過夜搖菌15h,加入等量60%甘油于-80℃超低溫保存待用。
本氏煙草轉(zhuǎn)化試驗參考王會強葉盤法轉(zhuǎn)化煙草的方法。切取無菌培養(yǎng)的煙草葉片預培養(yǎng)2d;農(nóng)桿菌侵染液濃度(OD600)為0.5,侵染時間為5~8min;煙草葉片與農(nóng)桿菌共培養(yǎng)最佳時間為2d。各時期培養(yǎng)基配方為:預培養(yǎng)、共培養(yǎng)培養(yǎng)基(MS+1mg/L6-BA+0.1mg/LNAA),分生培養(yǎng)基(MS+1mg/L6-BA+0.1mg/LNAA+50mg/LKana+500mg/LCef),生根培養(yǎng)基(MS+0.25mg/LNAA+50mg/LKana+500mg/LCef)。
2結(jié)果與分析
2.1HcMS1基因的克隆
TRIzol法提取RNA后取500ng進行逆轉(zhuǎn)錄得到cDNA用于后續(xù)試驗。以上述cDNA為模板擴增目的基因HcMS1,擴增產(chǎn)物瓊脂糖凝膠電泳檢測結(jié)果(圖1-A),擴增產(chǎn)物與目的片段大小一致且無雜帶。將擴增PCR產(chǎn)物切膠回收后克隆到T載體上,藍白斑篩選后進行菌落PCR擴增,擴增產(chǎn)物瓊脂糖凝膠電泳檢測結(jié)果(圖1-B)顯示,在2 000bp處有明顯的目的條帶,且無特異性擴增。分別取2個菌落PCR檢測為陽性的轉(zhuǎn)化子進行測序驗證,將測序結(jié)果與原始參考序列進行比對,HcMS1基因比對結(jié)果(圖2)顯示,測序結(jié)果和參考序列比對結(jié)果一致,克隆成功。

2.2HcMS1基因的生物信息學分析
利用Prot Param網(wǎng)站,對紅麻HcMS1基因編碼蛋白的理化性質(zhì)進行分析。結(jié)果顯示:HcMS1基因序列開放閱讀框(ORF)為1 950bp,共編碼氨基酸數(shù)649,包含20種常見氨基酸(表2),原子總數(shù)10 279,分子量73.566 72ku,分子式為C3270H5118N910O938S43,由于含有43個硫原子,故蛋白中可能存在二硫鍵。帶負電荷的殘基總數(shù)(Asp+Glu)為76,帶正電荷的殘基總數(shù)(Arg+Lys)為79,理論pI為7.89。HcMS1蛋白的不穩(wěn)定性指數(shù)為30.80,屬于穩(wěn)定蛋白質(zhì);脂肪指數(shù)為86.30,親水性指數(shù)平均值(GRAVY)為-0.221,更偏向為親水蛋白。

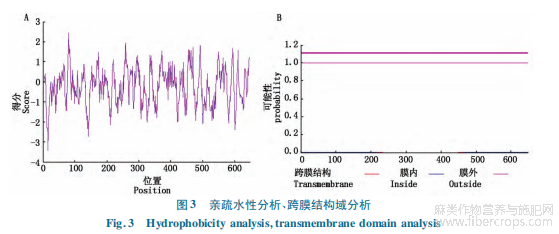
第16位氨基酸出現(xiàn)最低值-3.411,親水性最強,在第80位氨基酸出現(xiàn)最高值2.444,疏水性最強。整體看疏水氨基酸和親水氨基酸分布較均勻,偏向為親水蛋白。利用TMHMM在線分析HcMS1蛋白跨膜結(jié)構(gòu)域(圖3-B),HcMS1蛋白沒有跨膜結(jié)構(gòu)域,推測為膜外蛋白。
利用SOPMA預測HcMS1蛋白二級結(jié)構(gòu)(圖4-A),HcMS1蛋白由α-螺旋、延伸鏈、β轉(zhuǎn)角和無規(guī)則卷曲組成,分別占整個肽鏈的37.90%,15.87%,3.85%,42.37%。利用Phyre2模擬HcMS1蛋白三級結(jié)構(gòu)。建模過程中共搜索1 000條序列與目的蛋白序列進行比對,報告單按照置信度大小列出前120條序列模型,并以99.0%置信度,10%序列長度覆蓋度的d1weea(FYVE/PHD zincfinger)蛋白為參考建立模型(圖4-B)。
2.2.2信號肽、結(jié)構(gòu)域、亞細胞定位分析
通過SignalP-5.0Server分析(圖5-A),N端前70位氨基酸殘基,存在信號肽的概率幾乎為零,故推測Hc-MS1蛋白沒有信號肽,屬于非分泌蛋白。利用Inter-Pro在線分析(圖5-B),HcMS1蛋白預測到多個結(jié)構(gòu)域(591-641位Znf-PHD-finger、594-640位PHD、593-639位Znf-PHD),同源性超家族(590-644位Znf-FYVE-PHD、576-643位Znf-RING/FYVE/PHD)和保守序列(594-638位Zinc-finger-PHD-type-CS)。ProtComp9.0分析HcMS1蛋白亞細胞定位于細胞核。




2.2.3進化樹分析
在NCBI的BlastP中輸入HcMS1蛋白序列,在non-redundant protein sequences庫中搜索,獲得多個物種的同源蛋白并下載靠前的20個物種MS1蛋白序列,其中包括:陸地棉(Gos-sypium hirsutum)、榴蓮(Durio zibethinus)、木槿(Hi-biscus syriacus)、百花鶴望蘭(Herrania umbratica)、栓皮櫟(Quercus suber)、蓖麻(Ricinus communis)、巴西橡膠樹(Hevea brasiliensis)、番木瓜(Carica papaya)、毛果楊(Populus trichocarpa)、山茶花(Camelliasinensis)、桑樹(Morella rubra)、木薯(Manihot escu-lenta)、大麻(Cannabis sativa)、甜櫻桃(Prunus avi-um)、葡萄(Vitis vinifera)、相思子(Abrus precatori-us)、鷹嘴豆(Cicer arietinum)、月季(Rosa chinensis)、蒺藜苜蓿(Medicago truncatula)、大豆(Glycinemax)。利用MEGA7.0軟件,采用鄰近法構(gòu)建系統(tǒng)進化樹。分析構(gòu)建結(jié)果(圖6),其中陸地棉MS1蛋白與紅麻HcMS1蛋白進化關(guān)系最近,其次為木槿。
2.3HcMS1表達模式分析
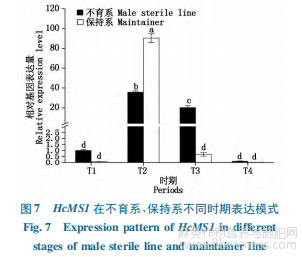
不同時期HcMS1表達模式分析顯示(圖7),在不育系和保持系的紅麻花藥中,HcMS1的表達隨著花蕾的發(fā)育均呈現(xiàn)先升高、后下降的趨勢,其在二期表達量最高,其次是三期,在四期的表達量最低,相對表達量僅為0.1,0.01。比較不育系與保持系發(fā)現(xiàn),保持系二期相對表達量為90.35,是不育系二期表達量的2.5倍,在P值0.05水平下呈現(xiàn)顯著差異。在保持系中,HcMS1于二期表達量超高,而在其他時期均為極低的表達水平,這與擬南芥MS1僅在小孢子釋放期表達事實相吻合;相比保持系,不育系HcMS1表達周期較長,在二期、三期均有相對較高的表達量,且在一期、四期表達水平均高于保持系。


2.4HcMS1基因過表達載體構(gòu)建和煙草轉(zhuǎn)化
2.4.1過表達載體構(gòu)建
將轉(zhuǎn)化的大腸桿菌進行PCR擴增,瓊脂糖凝膠電泳結(jié)果顯示,在2000bp處有明顯目的條帶(圖8)。取PCR陽性菌落提取質(zhì)粒后進行測序,比對結(jié)果顯示目的基因序列與參考序列一致,表示過表達載體構(gòu)建成功(圖9)。
2.4.2遺傳轉(zhuǎn)化
圖中培養(yǎng)基無污染、苗狀態(tài)健康(圖10-A)。將生根的煙草組培苗經(jīng)煉苗后移植到滅菌土中繼續(xù)培養(yǎng),提取野生型煙草和轉(zhuǎn)化煙草的新鮮葉片DNA,使用測序引物35S-F/2301-F(表1)進行PCR擴增片段,電泳后對比野生型發(fā)現(xiàn)轉(zhuǎn)化煙草在2000bp處有目的條帶,證明HcMS1轉(zhuǎn)化成功(圖10-B),對煙草成熟期觀察發(fā)現(xiàn):野生型煙草株型較高,可以正常結(jié)實,6片花萼大小均勻,花柱較長,果實大且飽滿;而轉(zhuǎn)基因煙草株型較矮,無法正常結(jié)實,6片花萼大小不一,花柱短小,兩者對比有明顯的差異(圖11)。表明通過葉盤法轉(zhuǎn)化獲得的HcMS1過表達轉(zhuǎn)基因本氏煙草無法正常自交結(jié)實,存在不育現(xiàn)象。
3討論與結(jié)論
紅麻不育機理研究目前主要在細胞生物學、分子生物學方面開展工作。在細胞生物學方面,主要通過研究花藥不同發(fā)育時期絨氈層和各組織化學成分變化,來確定花S藥敗育時期。大量研究表明,花藥絨氈層發(fā)育異常是花粉敗育的重要細胞學原因。朱麗梅等對紅麻細胞質(zhì)雄性不育系L23A和K03A的小孢子敗育過程進行了細胞學觀察,發(fā)現(xiàn)其敗育發(fā)生在四分體及單核小孢子時期。一年生麻類作物創(chuàng)新團隊在前期已通過對不育系LC0301A和保持系LC0301B不同時期的花藥細胞學觀察發(fā)現(xiàn),不育系發(fā)生敗育的主要時期為單核期,單核期不育系的絨氈層退化嚴重,小孢子出現(xiàn)畸形,不能形成正常的花粉粒。




MS1基因編碼PHD轉(zhuǎn)錄因子并參與調(diào)控花粉和絨氈層發(fā)育。目前MS1基因的功能在擬南芥中研究比較詳細,水稻、小麥等其他作物也相繼開展MS1基因的挖掘與分析工作。關(guān)于擬南芥的MS1基因的研究,早在21世紀初始便積極展開。Wilson等在2001年報道:擬南芥MS1突變體的花粉在四分體釋放小孢子后不久即發(fā)生退化,絨氈層也出現(xiàn)空泡化;MS1在小孢子釋放期的絨氈層部位低水平表達,缺失基因會導致其編碼蛋白的提前終止和PHD-finger基序的丟失。Ito等通過轉(zhuǎn)座子敲除擬南芥MS1基因,小孢子在四分體中分離出來即發(fā)生外層缺失;MS1基因僅在四分體的很短時期內(nèi)于絨氈層表達,并將MS1定位在細胞核上。2007年,Ito等通過構(gòu)建MS1-SRDX融合結(jié)構(gòu)轉(zhuǎn)化擬南芥揭示了MS1具有轉(zhuǎn)錄激活因子的功能,參與花粉外壁、花粉胞質(zhì)和絨氈層的發(fā)育。2006年,Vizcay-Barrena等通過TUNEL染色和超微結(jié)構(gòu)分析,野生型絨氈層在小孢子有絲分裂Ⅰ后發(fā)生程序性細胞死亡(PCD),而MS1突變體的絨氈層中未觀察到PCD的跡象。隨著人們認識到MS1基因?qū)φ{(diào)控擬南芥雄性不育的重要性,該基因的挖掘開發(fā)也在別的作物上相繼展開。2011年,Li等在水稻中的報道了一個擬南芥MS1同源基因PTC1,該基因編碼PHD-finger蛋白,控制絨氈層程序化發(fā)育和功能性花粉的形成,其突變體的絨氈層發(fā)生增殖失控和壞死樣死亡。2017年,Wang等在小麥中克隆并鑒定MS1基因,其在小孢子母細胞中特異性表達,編碼的蛋白定位于質(zhì)體和線粒體膜,具有磷脂酶結(jié)合活性;Tucker等發(fā)現(xiàn)小麥ms1可完全恢復了ms1d突變體的育性,編碼一種花粉外壁發(fā)育所必需的糖基磷脂酰肌醇錨定的脂質(zhì)轉(zhuǎn)移蛋白。本試驗從紅麻不育系和保持系的花藥中克隆出一個與擬南芥MS1基因相似度較大的同源基因HcMS1,前期通過細胞生物學的手段發(fā)現(xiàn)導致不育系花粉敗育的原因在于絨氈層PCD異常,絨氈層退化嚴重且提前,這與擬南芥中報道的MS1調(diào)控絨氈層PCD發(fā)育相似;同時通過qRT-PCR發(fā)現(xiàn)HcMS1花蕾二期的表達量在保持系中顯著高于不育系,且在保持系中該基因幾乎只在花蕾二期高度表達,這又與擬南芥MS1表達模式相符合,另外相比保持系,不育系在較長時間都保持著較高水平的表達,推測HcMS1表達時期的紊亂也可能是導致紅麻不育的原因之一。綜合以上,推測HcMS1基因在調(diào)控紅麻雄性不育的過程中亦發(fā)揮著一定的作用。
唐文武等以擬南芥花粉發(fā)育關(guān)鍵基因MS1為參考序列,通過Blast比對獲得蕓薹屬作物同源基因序列,運用生物信息學方法對其編碼氨基酸序列進行預測分析,為作物雜種優(yōu)勢利用提供理論參考。高嵩等同源克隆玉米ZmUdtl基因并利用生物信息學分析預測蛋白性質(zhì)、結(jié)構(gòu)等,為后期的克隆和功能鑒定奠定基礎(chǔ)。為了預測HcMS1基因的結(jié)構(gòu)功能,挖掘基因潛在信息,本試驗通過生物信息學方法對HcMS1蛋白進行預測分析。結(jié)果顯示,該蛋白為親水蛋白,沒有信號肽和跨膜結(jié)構(gòu),存在典型的PHD-finger結(jié)構(gòu)域,該結(jié)構(gòu)域廣泛存在于雄性不育基因MS1編碼蛋白中;其亞細胞定位在細胞核上,推斷HcMS1蛋白可能作為轉(zhuǎn)錄因子,作用于細胞核上,調(diào)控與育性相關(guān)的基因表達。進化樹分析結(jié)果顯示,紅麻HcMS1蛋白與陸地棉MS1蛋白親緣關(guān)系最近。
煙草作為一種模式植物,其轉(zhuǎn)基因體系已相當成熟,目前有很多在煙草上進行異源表達的研究,且大都獲得了與研究目的有關(guān)聯(lián)的價值性狀。試驗通過本氏煙草遺傳轉(zhuǎn)化發(fā)現(xiàn),35S強啟動子過表達HcMS1基因在陽性轉(zhuǎn)基因煙草中表現(xiàn)為植株矮化、花萼變小且大小不均、花柱縮短、無法結(jié)實,說明過表達HcMS1基因有影響本氏煙草正常育性的功能,為驗證HcMS1是調(diào)控紅麻雄性不育的關(guān)鍵基因提供了有力的支撐??傊?,本研究利用同源克隆方法在紅麻花藥中克隆出HcMS1基因,通過生物信息學分析發(fā)現(xiàn)HcMS1蛋白有顯著的PHD結(jié)構(gòu)保守域,其亞細胞定位于細胞核上,很有可能以轉(zhuǎn)錄因子的形式調(diào)控育性基因的表達。此外,qRT-PCR結(jié)果顯示,HcMS1基因在紅麻保持系花蕾的四分體至單核期高量表達,符合擬南芥MS1基因表達模式。最后,通過本氏煙草遺傳轉(zhuǎn)化試驗,在獲得的HcMS1過表達本氏煙草上發(fā)現(xiàn)了顯著的不結(jié)實現(xiàn)象。本研究克隆出紅麻HcMS1基因,并結(jié)合生物信息學、基因表達和煙草轉(zhuǎn)化對其進行分析,為挖掘紅麻不育調(diào)控基因提供了一定信息資源,為日后該基因的深度功能解析做好了鋪墊。
文章摘自 陳文濤,潘根,唐慧娟,常麗,李德芳,趙立寧,陳安國,李建軍.紅麻HcMS1基因的克隆、表達與功能分析[J].華北農(nóng)學報,2021,36(04):64-74.
更多閱讀
文章排行
最新文章
Copyright by Ramie Research Institute of Hunan Agricultural University
湖南農(nóng)業(yè)大學農(nóng)學院/苧麻研究所 版權(quán)所有 2009-2016 湘ICP備10006838號-1
