作者:彭文仙等
來(lái)源:
發(fā)布時(shí)間:2021-10-27
Tag:
點(diǎn)擊:
[麻進(jìn)展]7個(gè)苧麻AP2_ERF基因時(shí)空表達(dá)分析與花發(fā)育相關(guān)BnERF1基因克隆
摘 要:苧麻(Boehmeria nivea(L.)Gaudich.)生殖生長(zhǎng)對(duì)其纖維發(fā)育有顯著的抑制作用,且苧麻開(kāi)花受乙烯調(diào)控。AP2/ERF轉(zhuǎn)錄因子在植物中對(duì)乙烯信號(hào)響應(yīng)和調(diào)控生長(zhǎng)發(fā)育具有關(guān)鍵作用。本研究根據(jù)苧麻轉(zhuǎn)錄組數(shù)據(jù)篩選到7個(gè)不同的ERF基因,在分析其在不同種質(zhì)不同部位的時(shí)空表達(dá)情況基礎(chǔ)上,克隆了一個(gè)與花芽發(fā)育密切相關(guān)的ERF基因,并進(jìn)行生物信息學(xué)特征分析。結(jié)果表明:7個(gè)ERF基因在不同種質(zhì)和不同組織中均表達(dá),但表達(dá)量差異顯著,存在基因型和組織器官特異性。其中Unigene9156和Unigene16687在所有種質(zhì)中相對(duì)表達(dá)量較高,尤其在0.5~1cm花芽(雌花芽和雄花芽)中表達(dá)量極高。Unigene9156在‘GBN08’種質(zhì)0.5~1cm雄花芽中的表達(dá)量是在‘GBN08’麻骨中表達(dá)量的650多倍,而在‘GBN09’種質(zhì)0.5~1.0cm雌花芽的表達(dá)量是在‘GBN08’麻骨中表達(dá)量的338倍。因此可推測(cè)Unigene9156在調(diào)節(jié)苧麻開(kāi)花和花芽發(fā)育方面可能有重要作用。以苧麻轉(zhuǎn)錄組數(shù)據(jù)中的Unigene9156為模板,克隆unigene9156基因ORF長(zhǎng)為1302bp,命名為BnERF1(登錄號(hào):MZ540911)。該基因編碼433個(gè)氨基酸,分子質(zhì)量為47.05kD,理論P(yáng)I為9.04,含有AP2結(jié)構(gòu)域的YRG和RAYD基序,是無(wú)信號(hào)肽蛋白,亞細(xì)胞定位在細(xì)胞核內(nèi)。系統(tǒng)進(jìn)化樹(shù)顯示苧麻與川桑、異色山黃麻、大麻的ERF蛋白同源關(guān)系較近。該研究為進(jìn)一步揭示AP2/ERF調(diào)控苧麻開(kāi)花提供了科學(xué)依據(jù)。
關(guān)鍵詞:苧麻(Boehmeria nivea(L.)Gaudich.);AP2;ERF;克隆
苧麻(Boehmeria nivea L.Gaudich.)是以收獲韌皮纖維為主的多年生宿根性作物,三麻時(shí)期大量開(kāi)花結(jié)實(shí)顯示生殖生長(zhǎng)對(duì)苧麻纖維生長(zhǎng)有顯著的抑制作用,導(dǎo)致了纖維產(chǎn)量和品質(zhì)的下降(薛麗君,2015;馬鑫,2017)。苧麻開(kāi)花受乙烯調(diào)控。AP2/ERF(APETALA2與Ethylene-responsive factor)轉(zhuǎn)錄因子參與到乙烯信號(hào)的響應(yīng)與調(diào)控,在植物生長(zhǎng)發(fā)育等多個(gè)過(guò)程具有重要作用。開(kāi)展苧麻AP2/ERF轉(zhuǎn)錄因子研究可為分子水平調(diào)控苧麻開(kāi)花、減少或消除生殖生長(zhǎng),為生產(chǎn)中提供不開(kāi)花的苧麻品種或?yàn)橛N家提供不開(kāi)花的材料提供科學(xué)依據(jù)。
AP2/ERF轉(zhuǎn)錄因子是一個(gè)含有由60~70個(gè)氨基酸組成的AP2/ERF結(jié)構(gòu)域的轉(zhuǎn)錄因子家族,幾乎在所有的植物中都大量存在。通過(guò)內(nèi)部轉(zhuǎn)錄因子蛋白與其他相關(guān)蛋白之間的相互作用可以激活或抑制特定基因的轉(zhuǎn)錄。AP2/ERF轉(zhuǎn)錄因子一般含有DNA結(jié)合結(jié)構(gòu)域、轉(zhuǎn)錄激活或抑制結(jié)構(gòu)域、寡聚化位點(diǎn)以及核定位信號(hào)(吳慧敏等,2011)。根據(jù)AP2/ERF成員序列相似性和結(jié)構(gòu)域的數(shù)量,將其分為5類,分別為AP2、DREB、ERF、RAV和Soloist亞家族(Faraji et al.,2020)。ERF是植物AP2/ERF家族中的一個(gè)亞族。最先發(fā)現(xiàn)AP2基因與植物著花與種子發(fā)育相關(guān)。ERF蛋白質(zhì)的功能主要有以下幾個(gè)方面:植物防衛(wèi)反應(yīng)信號(hào)傳遞,應(yīng)答脅迫,調(diào)控基因表達(dá),調(diào)節(jié)植物的生長(zhǎng)發(fā)育等(許世達(dá)等,2021)。ERF家族基因的特征在于只含有1個(gè)AP2/ERF型DNA結(jié)合結(jié)構(gòu)域,能夠直接與乙烯響應(yīng)元件中的GCCbox相互作用,這對(duì)于乙烯調(diào)節(jié)轉(zhuǎn)錄過(guò)程十分重要(Xu et al.,2020)。前人有研究報(bào)道水稻OsERF71基因參與ABA信號(hào),并通過(guò)合成脯氨酸賦予植株抗旱性(Li et al.,2018);FaERF003可能作為乙烯響應(yīng)因子與乙烯調(diào)控草莓果實(shí)成熟密切相關(guān)(楊堉楠等,2021);在黏果酸漿中ERF基因調(diào)節(jié)花、果實(shí)的發(fā)育(鄭珍珍和黃蕓,2021)。據(jù)黃瓜RU性別研究相關(guān)報(bào)道,CsERF110和CsERF31具有分別結(jié)合A基因(CsACS11)和M基因(CsACS2)的啟動(dòng)子,激活其轉(zhuǎn)錄的功能(潘健等,2020),從而調(diào)控黃瓜花的性別(Tao et al.2018)。菊花的ERF基因轉(zhuǎn)入擬南芥中發(fā)現(xiàn)可以調(diào)控?cái)M南芥開(kāi)花時(shí)間(Xing et al.2019)。由此可見(jiàn)ERF基因的功能具有極大的物種特異性,但其作用機(jī)理尚不明晰,開(kāi)展不同作物中ERF基因的作用模式研究有重要的意義。
苧麻是雌雄同株異花植物,但雌雄花發(fā)育在不同時(shí)期,一般雄花先于雌花發(fā)育。前人研究發(fā)現(xiàn)雌性苧麻的乙烯釋放速率要大于雌雄同株苧麻;對(duì)于雌雄同株材料來(lái)說(shuō)雌花序的乙烯釋放速率>混合花序乙烯釋放速率>雄花序乙烯釋放速率;AVG和AgNO3都可以使苧麻明顯的雄性化,表明乙烯可以調(diào)節(jié)苧麻開(kāi)花,從而可以影響性別表達(dá)等。乙烯信號(hào)轉(zhuǎn)導(dǎo)研究發(fā)現(xiàn)(邢虎成等,2007),乙烯信號(hào)通過(guò)級(jí)聯(lián)反應(yīng)傳達(dá)給乙烯反應(yīng)元素結(jié)合蛋白(EREBP)轉(zhuǎn)錄因子基因(又名ERF),通過(guò)ERF的表達(dá)完成乙烯調(diào)控,調(diào)節(jié)最終應(yīng)答因子的反應(yīng),從而最終調(diào)控植物發(fā)育(張慧君,2013)。因此本研究基于前期苧麻錄組測(cè)序數(shù)據(jù)庫(kù),獲得了7個(gè)ERF家族基因,對(duì)其進(jìn)行不同苧麻種質(zhì)不同器官和不同發(fā)育時(shí)期的時(shí)空表達(dá)研究,從而確定與苧麻花發(fā)育密切相關(guān)的AP2/ERF轉(zhuǎn)錄因子基因,然后對(duì)其進(jìn)行克隆和生物信息學(xué)分析,為進(jìn)一步深入研究其的生物學(xué)功能提供科學(xué)依據(jù)。
1 結(jié)果與分析
1.1 7個(gè)ERF基因在旺盛生長(zhǎng)期的表達(dá)特性
基于湖南農(nóng)業(yè)大學(xué)邢虎成課題組與華大基因合作完成的苧麻開(kāi)花期轉(zhuǎn)錄組測(cè)序數(shù)據(jù)(采用solexa測(cè)序技術(shù)),我們得到了7條苧麻AP2/ERF片段序列。初步分析獲得序列,發(fā)現(xiàn)7條序列均含有AP2保守結(jié)構(gòu)域,說(shuō)明均屬于AP2/ERF轉(zhuǎn)錄因子家族。以這7條苧麻AP2/ERF片段序列為模板設(shè)計(jì)了實(shí)時(shí)定量PCR引物(表1),利用苧麻肌動(dòng)蛋白(Actin)基因作為內(nèi)參,采用表達(dá)量變化倍數(shù)對(duì)基因Unigene9156,Unigene18197,Unigene17478,Unigene16687,Unigene6896,Unigene15426,Unigene15247在5個(gè)苧麻品種不同器官間的表達(dá)情況進(jìn)行分析。在同一生長(zhǎng)時(shí)期不同的品種中這7個(gè)基因的表達(dá)量不同(圖A;圖B;圖C;圖D;圖E)。旺盛生長(zhǎng)期Unigene16687和Unigene9156基因在不同的苧麻種質(zhì)中均有表達(dá)。尤其在‘GBN08’苧麻種質(zhì)中,Unigene9156基因顯著高表達(dá)。其次則是Unigene16687基因,在‘圓青5號(hào)’中總相對(duì)表達(dá)超過(guò)了Unigene9156。7個(gè)苧麻AP2/ERF在骨、莖尖、皮、葉四個(gè)不同的組織中,也存在著組織表達(dá)特異性。Unigene16687和Unigene9156基因莖尖的表達(dá)較高,而Unigene18197和Unigene6896在5個(gè)不同品種的不同組織中表達(dá)量極低。Unigene17478,Unigene15426和Unigene15247在不同品種和組織部位中有表達(dá)情況,但三者的相對(duì)表達(dá)量都較低。由此可見(jiàn)Unigene9156和Unigene16687在苧麻中較為保守,有可能在生長(zhǎng)發(fā)育過(guò)程中承擔(dān)重要作用。
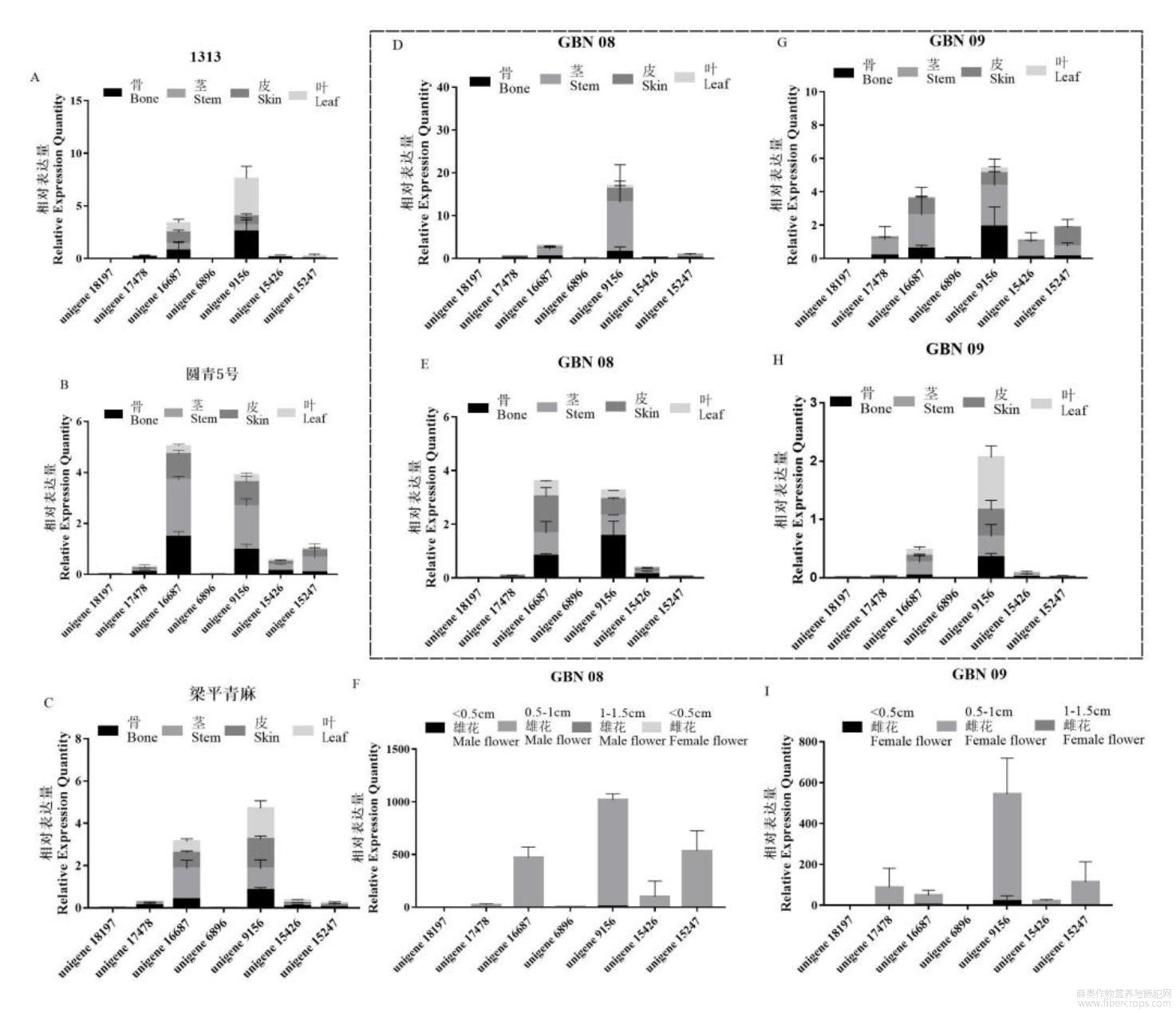
圖17個(gè)ERF基因在5個(gè)品種中的不同組織及不同品種不同生長(zhǎng)時(shí)期的表達(dá)特性
1.2 7個(gè)ERF基因在開(kāi)花期的表達(dá)特性
7個(gè)基因在開(kāi)花期的表達(dá)情況具有明顯差異,由于取樣時(shí)僅有‘GBN08’和‘GBN09’有花芽發(fā)育,因此試驗(yàn)觀察了‘GBN08’和‘GBN09’開(kāi)花期各不同組織7個(gè)ERF基因的表達(dá)情況。隨著苧麻生長(zhǎng),從旺盛生長(zhǎng)期到開(kāi)花期,7個(gè)ERF基因隨著苧麻的生長(zhǎng)發(fā)育在骨、莖尖、皮和葉組織中的表達(dá)量逐漸降低(圖1D,圖1F;圖1E,圖1G),而在花芽中的表達(dá)量極顯著的增加(圖1H;圖1I),尤其是Unigene9156和Unigene16687基因,在苧麻開(kāi)花期表達(dá)量顯著上調(diào),但Unigene16687的相對(duì)表達(dá)量上調(diào)倍數(shù)小于Unigene9156。無(wú)論是雌花還是雄花,雌雄同株的‘GBN08’或是全雌株的‘GBN09’中,Unigene9156基因幾乎都在花芽大小為0.5~1cm時(shí)高表達(dá),Unigene9156在‘GBN08’種質(zhì)0.5~1cm雄花芽中的表達(dá)量是在‘GBN08’麻骨中表達(dá)量的650多倍(圖1H,圖1I);該基因在‘GBN09’種質(zhì)0.5~1.0cm雌花芽的表達(dá)量是在‘GBN08’麻骨中表達(dá)量的338倍,在<0.5cm的雌花芽中表達(dá)量是在‘GBN08’麻骨中表達(dá)量的12倍。因此可推測(cè)Unigene9156在調(diào)節(jié)苧麻開(kāi)花和花芽發(fā)育方面可能有重要作用,可開(kāi)展進(jìn)一步研究。
1.3 花發(fā)育相關(guān)BnERF1基因ORF的克隆與生物信息學(xué)分析
時(shí)空表達(dá)分析發(fā)現(xiàn)Unigene9156可能與苧麻花芽發(fā)育有密切關(guān)系,因此以苧麻轉(zhuǎn)錄數(shù)據(jù)中的Unigene9156為模板,利用primer5.0設(shè)計(jì)基因克隆引物(表1),通過(guò)聚合酶鏈?zhǔn)椒磻?yīng)獲得克隆產(chǎn)物,最終得到約1300bp的條帶。純化后連接克隆載體轉(zhuǎn)入大腸桿菌,藍(lán)白斑選擇驗(yàn)證陽(yáng)性克隆,菌液測(cè)序結(jié)果分析表明苧麻Unigene9156基因ORF長(zhǎng)為1302bp,命名為BnERF1,(登錄號(hào):MZ540911)。該基因編碼433個(gè)氨基酸(圖2),該序列與轉(zhuǎn)錄組數(shù)據(jù)庫(kù)中的序列比對(duì)相似度達(dá)到99%以上。

圖2 苧麻BnERF1核苷酸序列及相應(yīng)氨基酸序列
通過(guò)在線軟件Expasy分析BnERF1基因的基本理化性質(zhì),其結(jié)果顯示該蛋白分子式為C2053H3235N595O651S12,分子質(zhì)量較小,僅為47.05kD,有利于后續(xù)研究;此蛋白是堿性蛋白質(zhì),理論P(yáng)I(等電點(diǎn))為9.04;不穩(wěn)定系數(shù)為50.72,脂肪系數(shù)為64.09,平均親水系數(shù)為:-0.706,負(fù)電荷殘基總數(shù)51。氨基酸組成中,G甘氨酸(Gly)和S絲氨酸(Ser)占比最高,二者都達(dá)到11.1%。
通過(guò)NCBI比對(duì)將BnERF1的ORF的氨基酸序列進(jìn)行結(jié)構(gòu)域分析,可以看出,其包含AP2的保守結(jié)構(gòu)域,符合AP2/ERF家族基因的特征。信號(hào)肽在線預(yù)測(cè)發(fā)現(xiàn)該蛋白無(wú)明顯信號(hào)肽??缒そY(jié)構(gòu)預(yù)測(cè)結(jié)果表明該蛋白沒(méi)有跨膜區(qū),不屬于跨膜蛋白。亞細(xì)胞定位在細(xì)胞核內(nèi),與其作為轉(zhuǎn)錄因子的作用位點(diǎn)相符。

圖3 不同物種ERF基因同源氨基酸比對(duì)結(jié)果

圖4不同物種中ERF氨基酸序列系統(tǒng)發(fā)育樹(shù)
苧麻BnERF1基因與其他15種植物ERF基因的氨基酸序列進(jìn)行同源性比對(duì),苧麻BnERF1基因含有AP2結(jié)構(gòu)域的YRG和RAYD基序,符合AP2/ERF家族基因的結(jié)構(gòu)特征(圖3)。系統(tǒng)進(jìn)化樹(shù)中苧麻與川(Morus notabilis,XP024027236.1)、異色山黃麻(Trema orientalis,PON86895.1)、大麻(Cannabis sativa,XP030495582.1)的ERF蛋白同源關(guān)系較近,與綠豆(Vigna radiata,XP014494817.1)、山核桃(Juglans regia,KAG2683513.1)、榴蓮(Durio zibethinus,XP022746128.1)等進(jìn)化關(guān)系較遠(yuǎn)(圖4)。經(jīng)以上序列分析發(fā)現(xiàn),BnERF1含有乙烯響應(yīng)因子的主要特征結(jié)構(gòu),提示可能在乙烯信號(hào)轉(zhuǎn)導(dǎo)或乙烯合成途徑中參與重要作用。
2 討論
苧麻作為一種飼纖兼用作物,具有纖維支數(shù)高、高蛋白、營(yíng)養(yǎng)豐富等特點(diǎn)。三麻時(shí)期開(kāi)花很大程度上影響了苧麻纖維品質(zhì)和營(yíng)養(yǎng)品質(zhì)。AP2/ERF基因家族是最大的植物特異性轉(zhuǎn)錄因子基因家族之一,在植物生長(zhǎng)發(fā)育、響應(yīng)外界脅迫以及系統(tǒng)進(jìn)化過(guò)程中發(fā)揮著重要作用,根據(jù)其AP2保守結(jié)構(gòu)域的特點(diǎn),分為了5個(gè)亞家族:ERF、DREB、AP2、RAV和soloist。含有一個(gè)AP2保守結(jié)構(gòu)域的是ERF亞家族與DREB亞家族,含有兩個(gè)重復(fù)的AP2結(jié)構(gòu)域的是AP2亞家族,而RAV亞家族成員則有含有兩個(gè)不同的結(jié)構(gòu)域,分別是一個(gè)AP2和一個(gè)B3結(jié)構(gòu)域,不同亞家族之間同源關(guān)系密切。目前有關(guān)ERF亞家族相關(guān)研究主要集中抗旱、抗?jié)澈涂果},如在小麥中(李世姣等,2021)分離出了4個(gè)鹽脅迫相關(guān)的ERF基因,包含脫落酸、水楊酸和茉莉酸等多種植物激素響應(yīng)元件,推測(cè)它們可能參與植物多種非生物脅迫信號(hào)轉(zhuǎn)導(dǎo)通路;瓦式獼猴桃(Bai et al.,2021)中AvERF73和AvERF78在耐澇基因型中特異性誘導(dǎo),而在敏感基因型中相對(duì)較低,表明它們?cè)谀蜐持械淖饔谩?Х榷?Matheus et al.,2021)中表明ERF基因可能作為體細(xì)胞胚性標(biāo)記,在植物形態(tài)建成與生長(zhǎng)發(fā)育中行使功能,這與本研究ERF基因在大部分苧麻發(fā)育期莖尖表達(dá)量較高的結(jié)果相吻合。Pan等(2021)的研究表明黃瓜CsERF31基因在乙烯信號(hào)通路上被CsEIN3激活,并刺激CsACS2從而觸發(fā)正反饋循環(huán)以確保雌性而非雙性花發(fā)育,CsERF31的敲除導(dǎo)致有缺陷的雙性花取代雌花。CsERF31的異位表達(dá)抑制雄花的雄蕊發(fā)育并促進(jìn)雌蕊發(fā)育,表明CsERF31具有性別轉(zhuǎn)換的功能因此CsERF31介導(dǎo)的正反饋回路在雌性黃瓜花發(fā)育過(guò)程中有重要意義。本研究中苧麻雌花和雄花的在0.5~1mm時(shí),BnERF1的相對(duì)表達(dá)量極顯著增加,或存在于黃瓜類似的調(diào)控機(jī)制,在雌蕊或雄蕊發(fā)育過(guò)程中有促進(jìn)或抑制作用,具體調(diào)控方式還有待進(jìn)一步的研究。Pei等(2021)人研究報(bào)道了棉屬(GoNe)的一個(gè)蜜腺發(fā)育基因,被注釋為APETALA2/ethylene-responsive factor。通過(guò)病毒誘導(dǎo)基因沉默技術(shù)和Cas9敲除的植物產(chǎn)生無(wú)蜜源表型。重復(fù)基因Ne1和Ne2同時(shí)點(diǎn)突變和基因截?cái)鄬?dǎo)致四倍體棉花蜜腺發(fā)育障礙,這也為苧麻開(kāi)花授粉的調(diào)控打開(kāi)了一個(gè)新思路,或許BnERF1基因也存在相似的功能。
3 材料與方法
3.1 植物材料
所有種質(zhì)種植于湖南農(nóng)業(yè)大學(xué)耘園基地,在苧麻旺盛生長(zhǎng)期取‘GBN08’(雌雄同株)、‘GBN09’(全雌株)、‘圓青5號(hào)’、‘1313’、‘梁平青麻’的骨、莖尖、皮和葉。在開(kāi)花期取‘GBN08’的花芽(<0.5cm雄花,0.5~1cm雄花,1~1.5cm雄花,<0.5cm雌花)、骨、莖尖、皮和葉,‘GBN09’的花芽(<0.5cm雌花,0.5~1cm雌花,1~1.5cm雌花)、骨、莖尖、皮和葉。其中‘GBN08’和‘GBN09’材料是湖南農(nóng)業(yè)大學(xué)苧麻遺傳育種課題組選育的不同性別的苧麻種質(zhì)。所有取樣樣品分離后用蒸餾水洗凈,用液氮迅速冷凍后于-80℃保存。
3.2 試驗(yàn)化學(xué)試劑
RNA提取試劑盒,cDNA合成試劑盒,購(gòu)自索萊寶生物公司。熒光定量PCR試劑盒購(gòu)自全式金(AQ141),其余試劑均為化學(xué)分析純。
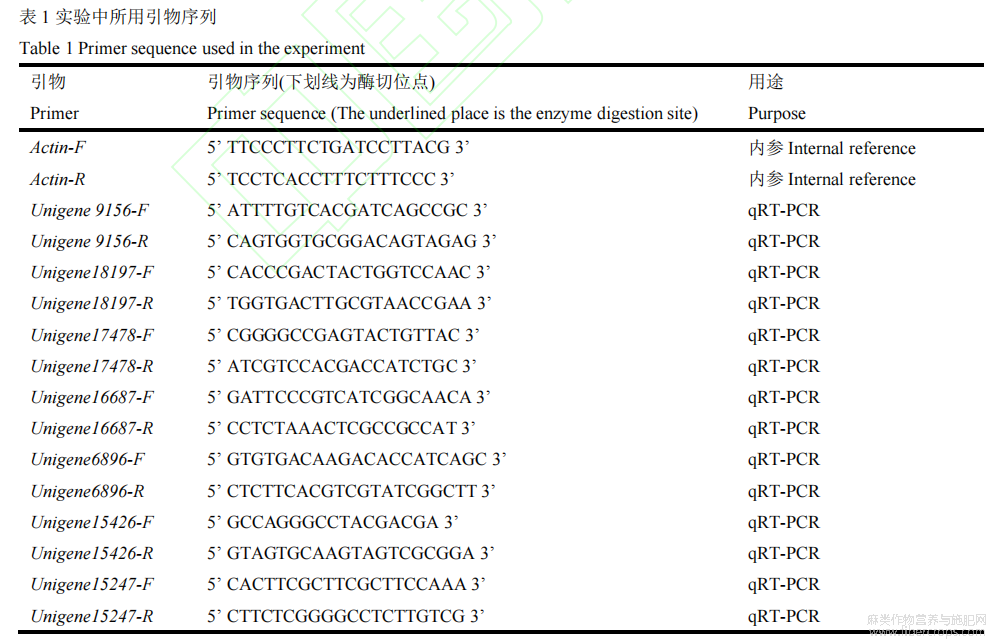
3.3 試驗(yàn)所用引物
以苧麻轉(zhuǎn)錄組序列為參考,生物信息學(xué)分析基因是否存在AP2結(jié)構(gòu)域,從而判斷是否為ERF家族成員,最終篩選到7個(gè)ERF家族成員,使用Primer5設(shè)計(jì)引物并由生工生物工程(上海)有限公司合成(表1)。
3.4 基因時(shí)空表達(dá)分析
試劑盒法提取材料中樣品的各組織RNA經(jīng)純度和完整性檢驗(yàn)后取1μg進(jìn)行反轉(zhuǎn)錄,反轉(zhuǎn)錄完成后的cDNA稀釋4倍用于實(shí)時(shí)熒光定量PCR實(shí)驗(yàn),反應(yīng)體系(20μL)如下:引物F0.4μL,引物R0.4μL,cDNA模板<1μg,Passive Reference Dye(50×)0.4μL,qPCRSuperMIX10μL,無(wú)酶水加至20μL。熒光PCR反應(yīng)程序:94℃變性3min,94℃變性30s,62℃退火30s,72℃延伸30s,40個(gè)循環(huán),72℃延伸10min。以苧麻的Actin作為內(nèi)參基因,數(shù)據(jù)處理使用2-℃℃ct的方法。
3.5 BnERF1基因的克隆
采用基因克隆引物進(jìn)行BnERF1基因的PCR克隆(表2),PCR反應(yīng)程序:98℃變性10s,55℃退火15s,72℃延伸2min30s,30個(gè)循環(huán)。PCR產(chǎn)物回收后p-EASY載體連接,構(gòu)建重組質(zhì)粒轉(zhuǎn)化大腸桿菌DH5α感受態(tài)細(xì)胞,通過(guò)藍(lán)白斑選擇進(jìn)行陽(yáng)性克隆驗(yàn)證后1:1甘油保存、測(cè)序。
3.6 BnERF1基因的生物信息學(xué)分析
使用DNAMAN軟件對(duì)測(cè)序回來(lái)的基因序列進(jìn)行拼接(NCBI登錄號(hào):MZ540911),在NCBI進(jìn)行核苷酸序列比對(duì)(NCBI網(wǎng)站Blast網(wǎng)址:https://blast.ncbi.nlm.nih.gov/Blast),并將推測(cè)的氨基酸序列進(jìn)行BlastP比對(duì)和結(jié)構(gòu)域預(yù)測(cè);二維結(jié)構(gòu)預(yù)測(cè)和二硫鍵預(yù)測(cè)(http://espript.ibcp.fr/ESPript/ESPript/);三維結(jié)構(gòu)預(yù)測(cè)(http://swissmodel.expasy.org);使用MEGA5.0軟件進(jìn)行氨基酸序列比對(duì)與系統(tǒng)發(fā)育進(jìn)化分析;蛋白質(zhì)基本理化性質(zhì)分析(http://web.expasy.org);信號(hào)肽預(yù)測(cè)(http://www.cbs.dtu.dk/services/SingalP/);跨膜結(jié)構(gòu)域預(yù)測(cè)(http://www.cbs.dtu.dk/services/TMHMM/);亞細(xì)胞定位預(yù)測(cè)(http://www.psort.org/。)
文章摘自:彭文仙,馬鑫,何思,何也君,張曉洋,霍穎怡,邢虎成.7個(gè)苧麻AP2/ERF基因時(shí)空表達(dá)分析與花發(fā)育相關(guān)BnERF1基因克隆[J/OL].分子植物育種:1-11[2021-10-20]
更多閱讀
文章排行
最新文章
Copyright by Ramie Research Institute of Hunan Agricultural University
湖南農(nóng)業(yè)大學(xué)農(nóng)學(xué)院/苧麻研究所 版權(quán)所有 2009-2016 湘ICP備10006838號(hào)-1
