作者:徐益等
來(lái)源:
發(fā)布時(shí)間:2021-10-12
Tag:
點(diǎn)擊:
[麻進(jìn)展] 主要麻類作物基因組學(xué)與遺傳改良:現(xiàn)狀與展望
摘 要:隨著測(cè)序技術(shù)的發(fā)展,主要麻類作物(黃麻、紅麻、苧麻、亞麻和工業(yè)大麻)參考基因組從 2011年至2020年陸續(xù)完測(cè)序,這標(biāo)志著麻類作物科學(xué)已經(jīng)進(jìn)入基因組時(shí)代。文章首先詳細(xì)概述主要麻類作物基因組測(cè)序。其次,評(píng)述了基于基因組學(xué)的麻類作物重要應(yīng)用價(jià)值基因挖掘?;趨⒖蓟蚪M和轉(zhuǎn)錄組測(cè)序,大量關(guān)于纖維發(fā)育、響應(yīng)非生物脅迫的候選基因被挖掘,以促進(jìn)麻類作物纖維的物種特性和“不與糧食爭(zhēng)好地”的逆境農(nóng)業(yè)。同時(shí)不同麻類作物特異性狀候選基因陸續(xù)被報(bào)道,如紅麻雄性不育、亞麻種子含油量和大麻大麻素相關(guān)候選基因等。再次,麻類作物基因組測(cè)序完成為基于組學(xué)的麻類作物遺傳改良提供可能:有助于麻類作物種質(zhì)資源形成和演化機(jī)制研究,系統(tǒng)解析纖維產(chǎn)量、纖維品質(zhì)、抗病耐逆等農(nóng)藝性狀形成的分子基礎(chǔ);有助于建立高通量基因型-表型數(shù)據(jù)庫(kù),挖掘優(yōu)異基因資源與創(chuàng)制新種質(zhì);有助于創(chuàng)新并集成分子標(biāo)記輔助選擇、基因組選擇、轉(zhuǎn)基因等技術(shù),建立高效的快速育種技術(shù)體系。宜選育高產(chǎn)高效、抗逆抗病、適宜輕簡(jiǎn)化機(jī)械化、優(yōu)質(zhì)專用的多用途麻類作物新品種,以滿足麻類作物相關(guān)產(chǎn)業(yè)的市場(chǎng)需求,適應(yīng)麻類作物生產(chǎn)方式。盡管已經(jīng)獲得重要基因以及位點(diǎn)的信息,但如何高效率利用已有基因資源對(duì)麻類作物進(jìn)行遺傳改良仍需面臨一系列挑戰(zhàn),如:成熟穩(wěn)定的遺傳轉(zhuǎn)化體系、麻類作物基因編輯體系構(gòu)建及基因組選擇育種等。
關(guān)鍵詞:主要麻類作物;基因組;基因;遺傳改良
黃麻、紅麻、苧麻、亞麻與工業(yè)大麻在種植歷史上曾經(jīng)超過(guò) 300 萬(wàn)公頃面積,屬于大面積栽培的麻類作物(以下簡(jiǎn)稱主要麻類作物),與糧、棉、油、菜并列為第 5 大作物群,其種植歷史可追溯到遠(yuǎn)古新石器至堯舜時(shí)代。主要麻類作物是重要的天然韌皮部纖維作物,其韌皮部纖維(麻皮)主要用于麻線、麻繩、麻袋、麻布等包裝用紡織,還用于家飾板材等多功能用途;其天然活性成分在食品、日用、醫(yī)藥等領(lǐng)域價(jià)值逐步開(kāi)發(fā),推動(dòng)生命健康產(chǎn)業(yè)發(fā)展。主要麻類作物分布范圍廣,在熱帶、亞熱帶、溫帶和寒帶均有分布。我國(guó)的華南、長(zhǎng)江流域、黃淮流域、淮河流域到東北(遼寧、吉林、黑龍江)、西北(新疆、寧夏、內(nèi)蒙古)等均可以種植。主要麻類作物生物學(xué)產(chǎn)量高,且耐鹽堿、耐干旱、耐洪澇、適應(yīng)性廣,不與糧、棉、油、菜爭(zhēng)好地,可用于鹽堿地或重金屬污染農(nóng)田的植物修復(fù),發(fā)展主要麻類作物將帶動(dòng)我國(guó)逆境農(nóng)業(yè)和麻類作物多功能利用的發(fā)展。
自1986年提出基因組學(xué)的概念,其相關(guān)研究不斷深入并被廣泛接受,目前基因組學(xué)已成為生命科學(xué)領(lǐng)域活躍且有影響的科學(xué)。模式植物擬南芥率先完成全基因組測(cè)序,這是第一個(gè)完成的植物基因組序列,隨后水稻也完成全基因組測(cè)序。隨著測(cè)序技術(shù)的發(fā)展,越來(lái)越多的植物進(jìn)行了全基因組測(cè)序,其中包括主要麻類作物(黃麻、紅麻、苧麻、亞麻和工業(yè)大麻)。這些主要麻類作物的全基因組測(cè)序,可以提供該作物全基因組的遺傳信息,尤其是對(duì)控制重要農(nóng)藝性狀的位點(diǎn)或基因的深入挖掘,以及對(duì)調(diào)控復(fù)雜性狀的分子網(wǎng)絡(luò)的解析奠定了重要基礎(chǔ)。
本文旨在總結(jié)主要麻類作物基因組學(xué)和遺傳改良研究的最新進(jìn)展,對(duì)于引領(lǐng)加快麻類作物遺傳育種研究具有重要意義。
1 主要麻類作物全基因組測(cè)序研究概況
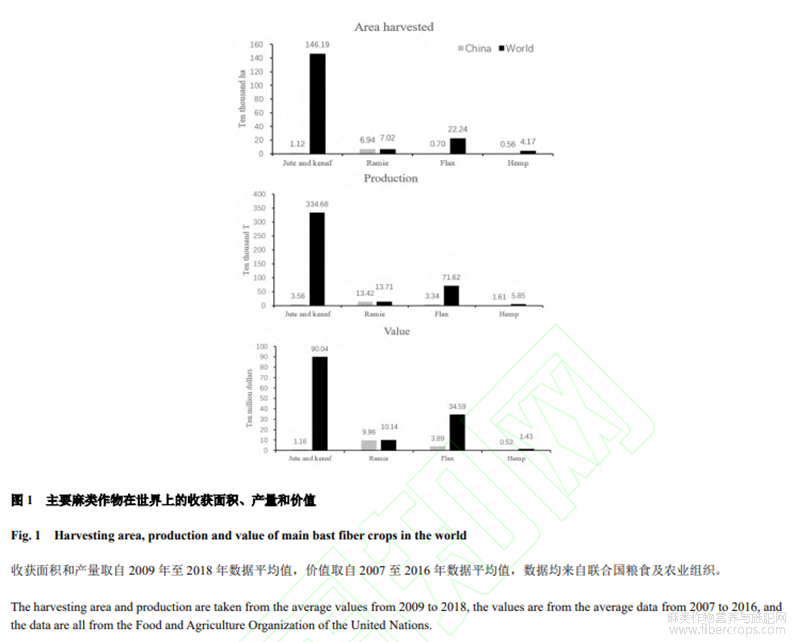
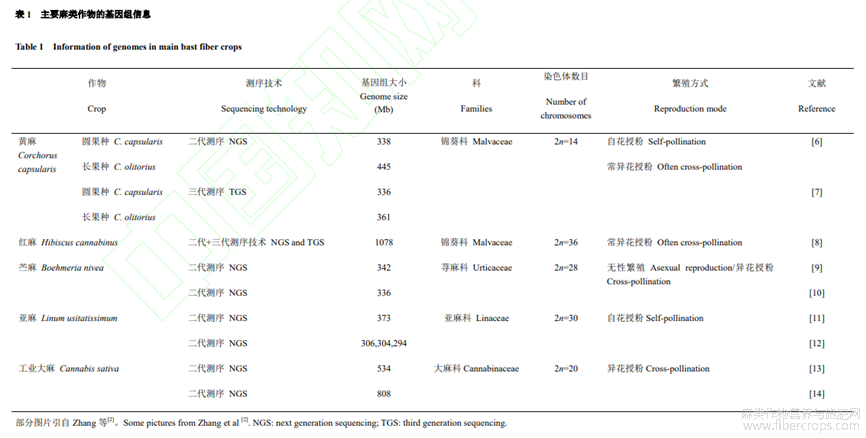
以聯(lián)合國(guó)糧食及農(nóng)業(yè)組織的 2007 年至 2018 年數(shù)據(jù)來(lái)看,在世界范圍內(nèi),黃麻和紅麻在收獲面積、產(chǎn)量和農(nóng)業(yè)生產(chǎn)價(jià)值均占據(jù)麻類作物之首,其次是亞麻、苧麻和工業(yè)大麻。其中我國(guó)黃麻和紅麻的收獲面積、產(chǎn)量和農(nóng)業(yè)生產(chǎn)價(jià)值居世界第三(圖1)。主要麻類作物基因組測(cè)序從 2011 年到 2020 年陸續(xù)公布,分別涉及錦葵科、蕁麻科、亞麻科和大麻科,基因組最小的是圓果種黃麻336 Mb,最大的是紅麻1078Mb(表1)。
1.1 黃麻基因組
黃麻基因組草圖于 2017 年發(fā)表,經(jīng)組裝發(fā)現(xiàn),長(zhǎng)果種 O-4 和圓果種 CVL-1 基因組大小分別為 445 Mb(13.04 Gb raw data,scaffold N50 為 3.3 Mb)和 338 Mb (13.69 Gb raw data,scaffold N50 為 4.1 Mb),分別有約8.4%和 17.8%的數(shù)據(jù)無(wú)法覆蓋,表明該研究只是得到了黃麻基因組草圖。為了獲得高質(zhì)量的黃麻參考基因組,福建農(nóng)林大學(xué)于 2015 年開(kāi)展黃麻轉(zhuǎn)錄組、基因組調(diào)研圖和流式細(xì)胞儀分析。在此基礎(chǔ)上,以黃麻國(guó)家區(qū)域試驗(yàn)對(duì)照品種長(zhǎng)果種寬葉長(zhǎng)果和圓果種黃麻179為材料,以三代測(cè)序技術(shù)PacBio RS II 為主體,Hi-C染色體構(gòu)象捕獲技術(shù)等技術(shù)輔助,獲得了長(zhǎng)果種和圓果種黃麻染色體級(jí)別的參考基因組,其基因組大小分別為361 Mb和336 Mb。與已發(fā)表的黃麻長(zhǎng)果種O-4和圓果種CVL-1 基因組相比,組裝指標(biāo)有了極大的提升。
黃麻全基因組的系統(tǒng)發(fā)育樹(shù)顯示,圓果種和長(zhǎng)果種黃麻均與可可和雷蒙德氏棉聚為一類,表明黃麻屬于錦葵科。需要指出的是,關(guān)于黃麻屬于錦葵科還是椴樹(shù)科一直以來(lái)存在爭(zhēng)議。為了驗(yàn)證這一結(jié)論,考慮到葉綠體表現(xiàn)為母系遺傳,其基因組被廣泛應(yīng)用于植物的系統(tǒng)發(fā)育關(guān)系分析,整理圓果種黃麻和長(zhǎng)果種黃麻以及紅麻的葉綠體基因組的系統(tǒng)發(fā)育關(guān)系結(jié)果發(fā)現(xiàn),黃麻圓果種和長(zhǎng)果種在親緣關(guān)系上與棉屬更近(附圖1)。這進(jìn)一步佐證黃麻屬于錦葵科,而不是基于植物形態(tài)學(xué)分類的椴樹(shù)科。
1.2 紅麻基因組
紅麻(Hibiscus cannabinus, 2n=36)是錦葵科木槿屬的一年生常異花授粉作物,屬于短日照喜溫類型。2020年福建農(nóng)林大學(xué)率先繪制高質(zhì)量紅麻參考基因組圖譜,該研究以紅麻優(yōu)良品種福紅952為材料,采用二代加三代的測(cè)序策略,同時(shí)結(jié)合Hi-C染色體構(gòu)象捕獲技術(shù)和高密度遺傳圖譜,完成紅麻全基因組測(cè)序和組裝工作,其基因組大小約為 1078 Mb,contig N50為2.73Mb,共鑒定到 66,004個(gè)基因。紅麻與擬南芥、水稻、亞麻和棉花的蛋白編碼基因比較發(fā)現(xiàn),2195個(gè)基因是紅麻所特有的,包括67個(gè)NAC和47個(gè)MYB轉(zhuǎn)錄因子。比較基因組學(xué)分析發(fā)現(xiàn),紅麻與雷蒙德氏棉在分化后存在一次獨(dú)立的全基因組復(fù)制事件(whole genome duplications, WGD),導(dǎo)致紅麻絕大多數(shù)基因拷貝數(shù)加倍。利用核心種質(zhì)重測(cè)序數(shù)據(jù),群體遺傳學(xué)分析顯示,紅麻起源于非洲,沿著非洲南部、非洲西部的路線傳播到亞洲,并在傳播過(guò)程中發(fā)生了2次馴化事件。在參考基因組基礎(chǔ)上,該研究確定了控制紅麻葉形的基因?yàn)長(zhǎng)ATE MERISTEM IDENTITY1 (LMI1)轉(zhuǎn)錄因子,并用病毒誘導(dǎo)的基因沉默(virus-induced gene silencing, VIGS)驗(yàn)證了其基因功能。
1.3 苧麻基因組
苧麻(Boehmeria nivea, 2n=28)為蕁麻科苧麻屬多年生宿根性草本植物,單性花,雌雄同株,花序復(fù)穗狀,雄花花序生在莖的中下部,雌花花序生于梢部。我國(guó)的苧麻產(chǎn)量占全世界苧麻產(chǎn)量的90%以上。
2018年2個(gè)苧麻基因組草圖陸續(xù)公布,以Zhongzhu 1為測(cè)序品種,得到一個(gè) 341.9 Mb的基因組草圖,contig N50和scaffold N50分別為22.62 kb和1226.36 kb,鑒定出30,237個(gè)蛋白編碼基因。苧麻作為蕁麻科中第一個(gè)被測(cè)序的物種,可對(duì)該物種的多樣性和物種形成等研究提供參考。比較基因組學(xué)發(fā)現(xiàn),苧麻有1075個(gè)獨(dú)有的基因家族,包含4082個(gè)基因,其中分別有5個(gè)和10個(gè)被注釋為假定的CesA和WAT1相關(guān)蛋白。分析這15個(gè)基因所編碼的蛋白結(jié)構(gòu)域發(fā)現(xiàn),5個(gè)假定 CesA 蛋白和3個(gè)假定WAT1 蛋白有相應(yīng)的保守結(jié)構(gòu)域,表明這些基因可能分別是真實(shí)的 CesA和WAT1相關(guān)基因,且與苧麻纖維產(chǎn)量有關(guān)。與其他4個(gè)物種(川桑、大棗、西瓜和亞麻)的 CesA和 WAT1相關(guān)蛋白的系統(tǒng)發(fā)育分析進(jìn)一步證實(shí),5個(gè) CesA和3個(gè) WAT1相關(guān)基因是苧麻特有的。進(jìn)化樹(shù)分析發(fā)現(xiàn)106個(gè)自然選擇的基因,其中22個(gè)與氮代謝有關(guān),這些基因可能與苧麻的粗蛋白含量和營(yíng)養(yǎng)生長(zhǎng)性狀馴化相關(guān)。這些研究為苧麻遺傳育種和分子研究提供了理論基礎(chǔ)。
1.4 亞麻基因組
2012年以CDC Bethune為測(cè)序品種,得到一個(gè)373 Mb的基因組草圖,contig N50為0.02 Mb,scaffoldN50為0.69 Mb,鑒定出43,484個(gè)基因。最近,甘肅農(nóng)科院分別對(duì)油用亞麻品種、纖維用亞麻品種和白胡麻測(cè)序得到染色體水平參考基因組,組裝得到的基因組大小為別為 306.0、303.7和293.5 Mb,其中contig N50/scaffold N50 分別為 131 kb/ 1235 kb、156 kb/700 kb 和 59 kb/384 kb,在每個(gè)基因組中大約預(yù)測(cè)到 43,500個(gè)編碼基因和2600~2800 個(gè)非編碼 RNA;系統(tǒng)發(fā)育分析表明,栽培亞麻和白胡麻在大約232萬(wàn)年前發(fā)生了分化,自古被子植物的六倍化事件以來(lái),亞麻發(fā)生了2次WGD事件。在最近一次WGD 事件中,MYB46/MYB83 基因拷貝數(shù)發(fā)生了擴(kuò)張,可能促成了亞麻獨(dú)特的次生細(xì)胞壁生物合成,對(duì) MYB46/MYB83基因的定向選擇可能形成了當(dāng)前的油用和纖維用亞麻的形態(tài)類型。對(duì)83份亞麻種質(zhì)資源重測(cè)序,在油用亞麻中,油脂相關(guān)基因(LOX、FatA、LTP 和 PDH-E2)和種子大小相關(guān)基因(BIN2和GW5)受到選擇;在纖維用亞麻中,次生細(xì)胞壁生物合成相關(guān)基因(MYB46-1、XTH和ROPGAP3)和植物株高相關(guān)基因(GA3ox、GA20ox和GID)受到選擇。
1.5 工業(yè)大麻基因組
2011年工業(yè)大麻基因組草圖發(fā)表,該研究以一種廣泛應(yīng)用于藥用的大麻品種 Purple Kush為測(cè)序品種,組裝得到一個(gè)534 Mb 的基因組草圖。2020 年 5 月公安部物證鑒定中心發(fā)表了一個(gè)高質(zhì)量染色體水平的野生大麻參考基因組,其基因組大小為 808 Mb,contig N50 為 513.57 kb,scaffold N50 為 83 Mb,相對(duì)于 2011 年發(fā)表的大麻基因組草圖(contig N50 為 2.8 kb;scaffold N50 為 16.2 kb)有很大的提升。統(tǒng)計(jì)其重復(fù)序列高達(dá) 74.75%,注釋到了 38,828 個(gè)蛋白編碼基因。然而目前為止籽用大麻基因組還未公布,待籽用大麻基因組公布之后,以期比較基因組學(xué)在 3 種不同用途的大麻之間挖掘更豐富的變異位點(diǎn),為不同用途的大麻育種提供更多信息。
2 主要麻類作物重要應(yīng)用價(jià)值基因挖掘
纖維發(fā)育與非生物脅迫相關(guān)基因的挖掘一直是主要麻類作物遺傳改良的重要方向。隨著主要麻類作物全基因組測(cè)序陸續(xù)被報(bào)道,很有必要系統(tǒng)總結(jié)回顧 NGS 技術(shù)對(duì)主要麻類作物纖維發(fā)育相關(guān)基因挖掘(附表1)、響應(yīng)各種非生物脅迫的遺傳機(jī)制(附表2)和麻類作物特異性狀候選基因的最新報(bào)道(附表3)。為了表述方便,不同麻類作物分開(kāi)闡述。每個(gè)麻類作物分別從纖維發(fā)育、非生物脅迫和麻類作物特異性狀3個(gè)方面展開(kāi)。
2.1 黃麻重要應(yīng)用價(jià)值基因挖掘
2.1.1 黃麻纖維發(fā)育相關(guān)基因
纖維發(fā)育的分子機(jī)制解析屬于麻類作物遺傳改良研究的重要方向。為鑒定纖維發(fā)育相關(guān)的基因,在黃麻纖維發(fā)育關(guān)鍵時(shí)期進(jìn)行轉(zhuǎn)錄組測(cè)序,共組裝出48,914 個(gè) unigenes,其中有5個(gè)Susy、3個(gè)UGPase、9個(gè)CesA、18個(gè)CSL、2個(gè)Kor (Korrigan),它們可能參與黃麻纖維素生物合成。長(zhǎng)果種黃麻莖皮轉(zhuǎn)錄組分析顯示,次生代謝產(chǎn)物的生物合成在代謝途徑中顯著富集,其中26個(gè)基因持續(xù)上調(diào),包括4個(gè)參與葡聚糖代謝的基因,調(diào)控莖皮韌皮部的發(fā)育。對(duì)一個(gè)纖維缺陷突變體進(jìn)行轉(zhuǎn)錄組測(cè)序發(fā)現(xiàn),CAD7在突變體的韌皮部組織中的任何生長(zhǎng)階段都顯著下調(diào),且CAD7在生長(zhǎng)早期的表達(dá)下調(diào)是伴隨著次生細(xì)胞壁特異性基因纖維素合成酶A7(CesA7)和阿拉伯半乳聚糖糖蛋白6 (FLA6)的共同上調(diào),這可能與黃麻纖維中協(xié)調(diào)木聚糖 S 型的沉積有關(guān)。黃麻全基因組測(cè)序顯示,在纖維素合成途徑中MYB83、WOX4、APL和HAT22的表達(dá)較高。抑制差減雜交被成功應(yīng)用于黃麻纖維發(fā)育過(guò)程中相關(guān)基因的鑒定,以纖維素含量極低的突變體作為試驗(yàn)組,纖維素含量正常的品種作為對(duì)照組,共鑒定出377個(gè)功能已知的基因。其中,COMT、CCR和4CL在正常植株中的表達(dá)明顯高于突變體,表明這些基因可能參與黃麻木質(zhì)素合成,且4CL是起主要作用。
2.1.2 黃麻非生物脅迫響應(yīng)基因
解析麻類作物非生物脅迫響應(yīng)的分子機(jī)制是其遺傳改良的又一重要方向,包括鹽脅迫和干旱脅迫等。黃麻鹽脅迫下轉(zhuǎn)錄組測(cè)序顯示,在葉和根中分別檢測(cè)到 32個(gè)和196個(gè)差異表達(dá)轉(zhuǎn)錄因子,其中有11個(gè)在2個(gè)組織中均被檢測(cè)到;在葉片中的差異表達(dá)轉(zhuǎn)錄因子家族包括 HB和 HSF,而在根中的差異表達(dá)轉(zhuǎn)錄因子家族包括 MYB、WRKY和CCAAT。脫落酸ABA 信號(hào)通路是植物鹽脅迫反應(yīng)的核心,在此途徑上發(fā)現(xiàn)了20個(gè)差異表達(dá)基因,包括編碼PP2C、SnRK2、ABF(上調(diào))和 PYL(下調(diào))的基因。在細(xì)胞分裂素途徑上發(fā)現(xiàn),在高濃度鹽脅迫下,葉和根中的 CRE1、B-ARR和A-ARR 均下調(diào),而大多數(shù) AHP 基因上調(diào),表明它們負(fù)調(diào)控細(xì)胞分裂素信號(hào)以提高植物的耐鹽性。黃麻耐旱品種和敏感品種的比較轉(zhuǎn)錄組分析,在干旱敏感型品種中,鑒定出34個(gè)轉(zhuǎn)錄因子差異表達(dá)基因和23個(gè)蛋白激酶差異表達(dá)基因,其中包括19個(gè)類受體激酶(receptor like kinases, RLKs)。這些 RLKs 在干旱脅迫下大部分下調(diào),說(shuō)明它們是黃麻抗旱機(jī)制的負(fù)調(diào)控因子。此外,3-ketoacyl-CoA合酶(3-ketoacyl-CoA synthase,KCS)的表達(dá)能顯著提高黃麻抗旱性,構(gòu)建 KCS 基因的過(guò)表達(dá)和基因編輯載體,并用于黃麻植株的轉(zhuǎn)化,發(fā)現(xiàn)KCS 基因的過(guò)表達(dá)不僅增加了葉綠體的數(shù)量,還增強(qiáng)了植株的抗旱性,提高了木質(zhì)素含量、生長(zhǎng)速率、莖皮厚度和莖粗。
2.1.3 黃麻特異性狀相關(guān)基因
黃麻2個(gè)栽培種(或亞種)在表型上存在較大差異,如:長(zhǎng)果種的果莢為長(zhǎng)條形狀,種子顏色為墨綠色;圓果種的果莢為球形,種子顏色為棕色等。不僅如此,長(zhǎng)果種為常異花授粉作物,圓果種為自花授粉作物,然而造成2個(gè)栽培種間形態(tài)或重要性狀差異的遺傳機(jī)制目前還有待深入研究。
2.2 紅麻重要應(yīng)用價(jià)值基因挖掘
2.2.1 紅麻纖維發(fā)育相關(guān)基因
與黃麻纖維不同的是紅麻韌皮纖維中纖維素含量相對(duì)較低,木質(zhì)素含量較高。紅麻基因組不僅揭示了一些參與韌皮纖維次生壁合成和纖維素沉積的基因,如:MYB、SND和CesA等,還結(jié)合QTL定位鑒定出MYB83和MYB103這2個(gè)可能參與細(xì)胞壁形成的基因,而MYB83也是紅麻在馴化過(guò)程中的受選擇基因。因此 MYB83 可能在紅麻纖維發(fā)育過(guò)程中發(fā)揮重要作用。轉(zhuǎn)錄組分析揭示出317個(gè)淀粉和糖代謝途徑的基因,可能參與纖維素生物合成。在此之前CAD、CCoAOMT、C3H、HCT、F5H和COMT被報(bào)道在木質(zhì)素生物合成中起關(guān)鍵作用。紅麻在開(kāi)花后纖維累積速度顯著降低,因此延后紅麻開(kāi)花期也是提高纖維產(chǎn)量的一個(gè)途徑,在紅麻光周期調(diào)控途徑中可能存在類似擬南芥 GI-CO-FT的保守途徑:短日照條件下,GI表達(dá)量上調(diào)促進(jìn)CO表達(dá),從而促進(jìn)植物開(kāi)花,GI和CO這2個(gè)基因在光敏感和光鈍感種質(zhì)中表達(dá)存在差異,說(shuō)明這2個(gè)基因可能參與紅麻光周期調(diào)控。
2.2.2 紅麻非生物脅迫響應(yīng)基因
紅麻本身作為一種耐旱、耐鹽堿、耐貧瘠、易栽培的作物,研究紅麻耐鹽堿機(jī)制,培育紅麻耐鹽堿種質(zhì)可將紅麻種植轉(zhuǎn)向鹽堿地,減少用地壓力。比較鹽脅迫下紅麻的轉(zhuǎn)錄譜發(fā)現(xiàn),鹽脅迫下轉(zhuǎn)錄因子NAC、BZIP、AP2/EREBP、ARF、AP2/ERF、bHLH、TCP均上調(diào),有8個(gè)NAC轉(zhuǎn)錄因子和2個(gè)WRKY轉(zhuǎn)錄因子表達(dá)下調(diào),這些轉(zhuǎn)錄因子可能通過(guò)多種不同途徑參與了紅麻的耐鹽性?;谵D(zhuǎn)錄組,不少學(xué)者對(duì)紅麻非生物脅迫產(chǎn)生響應(yīng)的基因進(jìn)入了深入挖掘,認(rèn)為WD40-1基因是紅麻ABA 和MeJA信號(hào)轉(zhuǎn)導(dǎo)途徑、鹽和干旱脅迫應(yīng)答途徑的關(guān)鍵樞紐基因;外源鎘和鹽處理能夠顯著誘導(dǎo)WRKY20表達(dá),該基因可能參與調(diào)控紅麻逆境脅迫反應(yīng),但仍需要更多試驗(yàn)驗(yàn)證 WRKY20 在紅麻逆境脅迫中的生物學(xué)功能。Wei 等分離鑒定紅麻6個(gè)HDACs 基因(HDA2、HHDA6、HDA8、HDA9、HDA19、SRT2),亞細(xì)胞定位顯示,HDA2 和 HDA8位于細(xì)胞核中,HDA6和HDA1位于細(xì)胞核和胞質(zhì)中,而HDA9位于細(xì)胞核和質(zhì)膜中,這6個(gè)基因均為紅麻鹽脅迫和干旱脅迫的響應(yīng)基因。
2.2.3 紅麻特異性狀相關(guān)基因
作物細(xì)胞質(zhì)雄性不育(cytoplasmic male sterile, CMS)是作物雜種優(yōu)勢(shì)利用的主要途徑,2004 年選育出紅麻 CMS 系 K03A,實(shí)現(xiàn)了三系配套,自此紅麻育性相關(guān)研究越來(lái)越多,與紅麻育性相關(guān)的基因也被陸續(xù)報(bào)道。趙艷紅等在紅麻線粒體基因ATP9中發(fā)現(xiàn)一個(gè)與雄性不育細(xì)胞質(zhì)相關(guān)的47bp缺失,這47 bp缺失片段分別與線粒體轉(zhuǎn)運(yùn)信號(hào)和GFP的ATP9融合,形成2個(gè)嵌合基因MTS-HM184-GFP和MTS-HM184,將含嵌合基因的載體轉(zhuǎn)入煙草發(fā)現(xiàn),部分轉(zhuǎn)基因植株為雄性不育后半不育,并基于ATP9 基因開(kāi)發(fā)的分子標(biāo)簽MM556可用于紅麻不育細(xì)胞質(zhì)的鑒定。對(duì)不育系和在保持系花藥進(jìn)行比較轉(zhuǎn)錄組分析發(fā)現(xiàn),ATP6在三羧酸循環(huán)(tricarboxylic acid cycle,TCA cycle)中起直接作用,在不育系中表達(dá)量下降了5.6倍。隨后ATP6基因也被報(bào)道因?yàn)槠銫DS中存在33-bp的缺失和 3-bp 的插入,導(dǎo)致紅麻細(xì)胞質(zhì)雄性不育。ATP6與 MYC2b蛋白之間存在互作,導(dǎo)致紅麻細(xì)胞質(zhì)雄性不育,而ATP6與MYC2a、MYC2b蛋白之間均存在互作時(shí),紅麻表現(xiàn)為可育。TIR1基因編碼一個(gè)富含亮氨酸重復(fù)的F-box蛋白,而F-bo蛋白被報(bào)道出在花器官發(fā)育中有重要功能,且在花藥期不育系中TIR1基因表達(dá)顯著高于保持系,這可能導(dǎo)致花藥敗育。目前認(rèn)為雄性不育主要包括由線粒體基因和核基因共同控制,然而又有報(bào)道通過(guò)RNA編輯技術(shù)發(fā)現(xiàn)葉綠體基因也可能與紅麻細(xì)胞質(zhì)雄性不育有關(guān)。紅麻細(xì)胞質(zhì)雄性不育基因及其調(diào)控機(jī)制還有待深入研究。
2.3 苧麻重要應(yīng)用價(jià)值基因挖掘
2.3.1 苧麻纖維發(fā)育相關(guān)基因
苧麻纖維中纖維素含量高于黃麻且木質(zhì)素含量遠(yuǎn)低于黃麻,纖維品質(zhì)比黃麻好。轉(zhuǎn)錄組測(cè)序鑒定出36個(gè)纖維素合酶基因,其中有33個(gè)在莖皮中的表達(dá)遠(yuǎn)高于其他組織。隨后CesA1、CesA2、CesA3和 CesA4也都被陸續(xù)報(bào)道在韌皮部或木質(zhì)部表達(dá)參與次生細(xì)胞壁合成。對(duì)苧麻野生種和栽培種的轉(zhuǎn)錄組進(jìn)行比較發(fā)現(xiàn),高纖維素含量品種的馴化可能與WAT1 基因的正向選擇有關(guān)。WAT1相關(guān)的基因在苧麻基因組中也被注釋為與纖維產(chǎn)量的相關(guān)基因。對(duì)木質(zhì)素含量的遺傳基礎(chǔ)進(jìn)行分析發(fā)現(xiàn),3個(gè)位于木質(zhì)素含量 QTL 區(qū)域的基因,在纖維發(fā)育的2個(gè)不同階段都有不同的表達(dá),且它們分別編碼 MYB 蛋白、莽草酸羥基肉桂酰轉(zhuǎn)移酶和漆酶。Tang 等發(fā)現(xiàn),PAL、C4H、4CL1、F5H、CCAOM和CAD的表達(dá)量均與木質(zhì)素含量成正比,但苧麻4CL3重組蛋白在催化過(guò)程中以肉桂酸為底物,因此對(duì)木質(zhì)素合成有負(fù)調(diào)控作用,推測(cè)苧麻4CL3主要參與苧麻黃酮類化合物的合成,4CL1主要參與木質(zhì)素的合成。探索 NAC 與 MYB 對(duì)調(diào)控苧麻纖維次生細(xì)胞壁發(fā)育的作用發(fā)現(xiàn),NAC19 和 NAC24 過(guò)表達(dá)能夠上調(diào)苧麻次生壁發(fā)育關(guān)鍵基因 MYB46 的表達(dá)量,從而導(dǎo)致次生壁纖維的生物合成增加;NAC19 有可能直接參與細(xì)胞程序性死亡調(diào)控次生細(xì)胞壁發(fā)育,而 MYB46 可能在苧麻纖維成熟中發(fā)揮重要作用。對(duì)不同區(qū)段的莖皮進(jìn)行轉(zhuǎn)錄組測(cè)序發(fā)現(xiàn),乙烯和赤霉素可能參與的苧麻韌皮纖維組織發(fā)育,這說(shuō)明研究調(diào)控苧麻纖維發(fā)育的相關(guān)基因可以間接從乙烯和赤霉素合成酶入手。
2.3.2 苧麻非生物脅迫響應(yīng)基因
非生物脅迫嚴(yán)重影響苧麻的產(chǎn)量與品質(zhì)?;谄r麻轉(zhuǎn)錄組,篩選出 8個(gè)WRKY轉(zhuǎn)錄因子,與其他作物已報(bào)道的10個(gè)具有抗旱功能的WRKY轉(zhuǎn)錄因子對(duì)比,推斷這個(gè)8個(gè)WRKY轉(zhuǎn)錄因子也可能參與苧麻干旱脅迫響應(yīng)。ACO1、bZIP1和 α-amylase也被證實(shí)響應(yīng)苧麻 ABA、干旱和高鹽逆境脅迫,將苧麻 GS2 基因轉(zhuǎn)入煙草發(fā)現(xiàn),其顯著提高轉(zhuǎn)基因植株的生物量和氮利用效率,從而減輕外界非生物脅迫。PCS1、MYB83、G6PDH1和 NRAMP1也都被陸續(xù)報(bào)道出能響應(yīng)重金屬鎘脅迫。此外,Chen 等發(fā)現(xiàn) 73 個(gè) miRNAs,它們介導(dǎo)苧麻對(duì)重金屬鎘脅迫的響應(yīng)。
2.3.3 苧麻特異性狀相關(guān)基因
苧麻是一個(gè)雜種優(yōu)勢(shì)利用率較高的作物,對(duì)苧麻育性相關(guān)基因的挖掘能為其雄性不育機(jī)理研究奠定基礎(chǔ)。目前已經(jīng)報(bào)道出的與苧麻育性相關(guān)的基因有 ATPA、ATP6和ATP9。
2.4 亞麻重要應(yīng)用價(jià)值基因挖掘
2.4.1 亞麻纖維發(fā)育相關(guān)基因
亞麻纖維中纖維素含量高于黃麻,木質(zhì)素含量遠(yuǎn)低于黃麻,纖維品質(zhì)優(yōu)于黃麻。基于亞麻基因組信息,鑒定出 32 個(gè)與亞麻纖維發(fā)育相關(guān)的候選基因,其中 16 個(gè)編碼纖維素合成酶(CesA),16 個(gè)編碼纖維素合成酶類蛋白(Csl),利用 VIGS 沉默這些基因后,韌皮纖維的數(shù)量和結(jié)構(gòu)受到了嚴(yán)重的影響。亞麻纖維素生物合成的調(diào)節(jié)涉及CesA的表達(dá)調(diào)控、胞質(zhì)中纖維素合酶復(fù)合體的裝配、纖維素的沉積等多個(gè)方面,但對(duì)Csl蛋白的功能尚不清楚,而 CesA 基因的轉(zhuǎn)錄水平主要依賴于產(chǎn)生的蛋白質(zhì)的功能亞類和植物發(fā)育階段。對(duì)亞麻不同莖段的轉(zhuǎn)錄組分析顯示,在次生細(xì)胞壁形成的晚期韌皮纖維中(莖底部區(qū)域),編碼金屬硫蛋白、脂質(zhì)轉(zhuǎn)移蛋白以及參與蛋白質(zhì)合成/翻譯和泛素介導(dǎo)降解的候選基因表達(dá)量增加;與頂桿區(qū)相比,分化后期的韌皮纖維中有156個(gè)基因上調(diào),其中有與細(xì)胞壁相關(guān)的基因;與其他莖段相比,參與次生細(xì)胞壁沉積的轉(zhuǎn)錄因子在韌皮纖維中的表達(dá)減少。除以上基因家族外,COMT基因主要參與S-木質(zhì)素的生物合成,轉(zhuǎn)基因植物在 COMT活性被抑制時(shí),木質(zhì)素含量減少;PHAC1 基因表達(dá)量與亞麻纖維拉力強(qiáng)度成正比[68];Ca2+信號(hào)通路通過(guò)對(duì) PLR1 基因的轉(zhuǎn)錄調(diào)控,在木質(zhì)素生物合成的 ABA 調(diào)控中起著至關(guān)重要的作用,而 CML15b 是該調(diào)控的關(guān)鍵調(diào)節(jié)因子;WRKY36 通過(guò)與 PLR1 啟動(dòng)子中的 W-box 結(jié)合,在響應(yīng) ABA 和尖孢鐮刀菌誘導(dǎo)的木質(zhì)素生物合成中發(fā)揮調(diào)節(jié)作用。
2.4.2 亞麻非生物脅迫響應(yīng)基因
亞麻基因組測(cè)序完成為全基因組水平分析非生物脅迫響應(yīng)基因提供了前提條件。在亞麻全基因組中共找到 137個(gè)NBS類抗病基因,34個(gè)HSF基因響應(yīng)高溫脅迫。轉(zhuǎn)錄組測(cè)序發(fā)現(xiàn),WRKY和JAS基因家族在亞麻對(duì)土壤營(yíng)養(yǎng)脅迫的響應(yīng)中起著重要的作用,CAX3基因產(chǎn)物可能通過(guò)Ca2+介導(dǎo)的胞內(nèi)調(diào)節(jié)參與了亞麻對(duì)高酸性、高Al3+濃度的響應(yīng),MADS-box和 NAC轉(zhuǎn)錄因子參與調(diào)節(jié)植物生長(zhǎng)發(fā)育和參與亞麻耐鋁性細(xì)胞壁修飾的酶。干旱脅迫對(duì)亞麻產(chǎn)量和油質(zhì)量的影響主要表現(xiàn)在生育期,選擇性掃描發(fā)現(xiàn)了與脫落酸途徑、生長(zhǎng)素信號(hào)、Ca2+信號(hào)、光合作用調(diào)節(jié)和干旱應(yīng)答轉(zhuǎn)錄因子有關(guān)的各種基因。亞麻銹病感染轉(zhuǎn)錄組揭示6個(gè) Avr 基因(AvrM、AvrM14、AvrL2、AvrL567、AvrP123(AvrP)和 AvrP4)參與了亞麻抵御銹病感染,在感染早期表達(dá)量達(dá)到一個(gè)峰值,隨著孢子形成逐漸下降。
2.4.3 亞麻特異性狀相關(guān)基因
籽用亞麻種子含油量一般為30%~45%,早在史前時(shí)代就被作為油料作物種植。對(duì)亞麻未成熟胚中磷脂酶基因家族的轉(zhuǎn)錄組學(xué)分析發(fā)現(xiàn),PLA2在亞麻中主要參與甘油磷脂代謝途徑和醚酯代謝途徑;PLC除參與甘油磷脂代謝和醚酯代謝之外,還在肌醇磷酸代謝途徑中發(fā)揮重要作用。在TAG合成途徑中發(fā)現(xiàn)7個(gè)關(guān)鍵基因,其中 PDAT1、DGAT1和 DGAT2基因的動(dòng)態(tài)表達(dá)模式與含油量的動(dòng)態(tài)積累模式顯著正相關(guān),PDAT1的動(dòng)態(tài)表達(dá)模式與亞麻酸的動(dòng)態(tài)積累模式顯著正相關(guān),且在高油高亞麻酸材料的種子動(dòng)態(tài)發(fā)育階段中 PDAT1基因的累積表達(dá)量也顯著高于低油低亞麻酸材料,因此 PDAT可能是影響籽用亞麻(胡麻)不同品種(系)中含油量和亞麻酸含量的關(guān)鍵基因。全基因組關(guān)聯(lián)分析和結(jié)合轉(zhuǎn)錄組測(cè)序,篩選出10個(gè)候選基因,其中6個(gè)基因參與了重要的脂肪酸代謝途徑,其中一些基因還具有上下游調(diào)節(jié)關(guān)系。對(duì)不同亞麻酸含量的亞麻品種進(jìn)行不同時(shí)期的品質(zhì)測(cè)定發(fā)現(xiàn),F(xiàn)AD2a、FAD3a 和 FAD3b這3個(gè)基因參與不飽和脂肪酸積累過(guò)程。
2.5 工業(yè)大麻重要應(yīng)用價(jià)值基因挖掘
2.5.1 工業(yè)大麻纖維發(fā)育相關(guān)基因
工業(yè)大麻是一種多用途作物,韌皮纖維主要用于紡織和生物復(fù)合工業(yè)。其下胚軸有一個(gè)主動(dòng)伸長(zhǎng),然后變厚形成初生和次生韌皮纖維的階段,因此工業(yè)大麻是研究二次生長(zhǎng)過(guò)程的合適模型。工業(yè)大麻下胚軸的二次生長(zhǎng)與轉(zhuǎn)錄因子 NST1、MYB46 和 WLIM1 的上調(diào)有關(guān)。韌皮纖維的伸長(zhǎng)依賴于 XTH 的活性,如 XTH5 和 XTH8,這些基因在伸長(zhǎng)的大麻下胚軸中表達(dá)較多。COBRA 家族中參與纖維素生物合成的 2 個(gè)基因 COB 和 COBL4,以及次生細(xì)胞壁纖維素合成酶基因(CesA4、7 和 8)在二次生長(zhǎng)的下胚軸中均有較高表達(dá)量。FLA11 和 FLA12 在工業(yè)大麻二次生長(zhǎng)的下胚軸中表達(dá)量高,此時(shí)初生和次生韌皮纖維都存在。已知 FLA 基因家族會(huì)影響植物細(xì)胞壁中纖維素、阿拉伯糖和半乳糖的含量,因此也可能有助于確定韌皮纖維細(xì)胞壁的組成。RNA-Seq 分析發(fā)現(xiàn),在下胚軸二次生長(zhǎng)過(guò)程中涉及到葡萄糖醛酸苷生物合成的幾個(gè)基因的上調(diào),特別是涉及到主干延伸(IRX10、IRX10L、IRX14和 IRX15L)、主干乙酰化(ESK1、RWA3)和取代基甲基化(GXM3)有關(guān)的基因。
2.5.2 工業(yè)大麻特異性狀相關(guān)基因
工業(yè)大麻雖然是一種纖維作物,但大麻素作為工業(yè)大麻所特有的一類次生代謝產(chǎn)物,在抗腫瘤、神經(jīng)系統(tǒng)保護(hù)、免疫調(diào)節(jié)和抗炎抗氧化等方面具有重要的藥用價(jià)值,因此工業(yè)大麻作為藥用植物也有很大的前景。工業(yè)大麻全基因組測(cè)序鑒定出2個(gè)參與調(diào)控大麻素合成途徑的基因 AEE1 和 OLS,此外還鑒定出23 個(gè)編碼大麻素原酸(CBCA)形成的候選基因,其中4個(gè)基因命名為THCAS-like1~THCAS-like4。在 MEP 和 GPP 途徑后端的酶基因 CMK、MDS、HDS、HDR和GPP (lsu)表達(dá)量和大麻素含量呈現(xiàn)顯著正相關(guān),在大麻素合成途徑中OLS、PT和THCAS 3 個(gè)大麻中特有的酶基因主要在始果期苞片腺毛中對(duì)大麻素合成積累起著關(guān)鍵作用。不同工業(yè)大麻品種的腺毛轉(zhuǎn)錄組分析,鑒定出之前從未報(bào)道過(guò)的編碼活性酶的基因,TPS18VF和 TPS19BL編碼橙花醇/芳樟醇合成酶,TPS16CC 編碼大根香葉烯B合酶和TPS20CT編碼四甲基環(huán)癸二烯甲醇合酶。大麻聚酮合酶(PKS)是催化大麻素生物合成的第一步,有研究發(fā)現(xiàn),啤酒花衍生的轉(zhuǎn)錄因子和番茄濃密矮化病毒(TBSV)p19 共同參與了PKS 基因啟動(dòng)子的激活。Laverty等利用藥用大麻和毒品大麻雜交群體構(gòu)建遺傳圖譜發(fā)現(xiàn),大麻素的生物合成基因是不連鎖的,但產(chǎn)生THCA和CBDA合成酶底物的芳香族丙烯酰胺轉(zhuǎn)移酶(AP)與已知的大麻素含量標(biāo)記緊密連鎖,這為在大麻中如何產(chǎn)生大麻素提供了不同見(jiàn)解。
大麻作為雌雄異株植物,性染色體進(jìn)化是其又一特異性狀。Prentout等在大麻中發(fā)現(xiàn)了超過(guò)500個(gè)性連鎖基因,將這些性連鎖基因定位到大麻基因組組合中,其中有363個(gè)在1號(hào)染色體,并且發(fā)現(xiàn)有1對(duì)染色體具有較大的非重組區(qū),進(jìn)一步分析發(fā)現(xiàn),大麻具有一個(gè)強(qiáng)烈退化的Y染色體,是迄今為止最古老的植物性染色體系統(tǒng),這有助于工業(yè)大麻性染色體鑒定和性別基因定位。
3 基于基因組學(xué)的麻類作物遺傳改良
3.1 我國(guó)麻類作物品種改良?xì)v程
主要麻類作物在我國(guó)均擁有悠久的種植歷史,其品種改良大致經(jīng)歷了以下4 個(gè)階段:第1階段是地方種的搜集鑒定及引種利用。在此階段不僅大量豐富了種質(zhì)資源類型,而且鑒定出一批具有優(yōu)良性狀的材料,如:黃麻品種D154和翠綠等。第2階段是雜交育種或回交選育等常規(guī)育種方法選育優(yōu)良種質(zhì)。此階段是利用第 1 階段所得到的優(yōu)異種質(zhì)為基礎(chǔ)材料進(jìn)行育種工作,在此階段利用雜交與誘變育種相結(jié)合,選育出一大批高纖維產(chǎn)量和高含油量的亞麻品種,如:黑亞 2 號(hào)和亞寧 1 號(hào)等。第 3 階段為高產(chǎn)、優(yōu)質(zhì)、多抗育種階段。在此之前由于長(zhǎng)期使用少數(shù)幾個(gè)性狀優(yōu)異的骨干材料作為親本,使麻類作物品種遺傳基礎(chǔ)狹窄,導(dǎo)致作物平均單產(chǎn)和品質(zhì)的降低以及病蟲(chóng)害的發(fā)生,如:亞麻枯萎病和紅麻炭疽病,因此育種家們?cè)诖穗A段選育出一批高產(chǎn)、優(yōu)質(zhì)、多抗育種的新品種。第 4 階段為優(yōu)質(zhì)高產(chǎn)專用新品種選育和現(xiàn)代生物技術(shù)輔助育種階段。由于常規(guī)育種在進(jìn)一步提高產(chǎn)量、品質(zhì)和抗性等方面的局限性逐步顯現(xiàn),且很難滿足市場(chǎng)需求的優(yōu)質(zhì)專用種質(zhì)培育,與此同時(shí)現(xiàn)代生物技術(shù)快速發(fā)展,利用生物技術(shù)輔助育種越來(lái)越受到重視,因此麻類作物已經(jīng)由單一的纖維用新品種選育向多用途、專用型新品種選育發(fā)展,如:亞麻輪選1號(hào)和天亞7號(hào)等,紅麻福紅2號(hào)和福紅992等??梢?jiàn),我國(guó)主要麻類作物的育種方法正在從常規(guī)育種轉(zhuǎn)變?yōu)槌R?guī)育種結(jié)合現(xiàn)代生物技術(shù)輔助育種方法,而育種目標(biāo)也在從高產(chǎn)轉(zhuǎn)變?yōu)閮?yōu)質(zhì)抗病高產(chǎn)并重,逐步過(guò)渡到為了適用多用途而進(jìn)行專用型品種的選育。其遺傳改良已經(jīng)從一個(gè)完全基于表型的過(guò)程,在某種程度上轉(zhuǎn)變?yōu)樵絹?lái)越依賴基于基因型的選擇。這得益于基因組學(xué)革命極大地提高了我們對(duì)作物基因組的理解,使得現(xiàn)代育種家可以更加關(guān)注一個(gè)新的時(shí)代,即基于基因組測(cè)序的作物遺傳改良。
3.2 麻類作物育種技術(shù)的變遷
我國(guó)麻類作物育種工作開(kāi)始于農(nóng)家品種選育及外來(lái)優(yōu)異種質(zhì)引進(jìn)結(jié)合常規(guī)育種手段,如雜交育種、誘變育種等,選育出性狀優(yōu)良種質(zhì)。隨著生物技術(shù)的高速發(fā)展,常規(guī)育種手段結(jié)合現(xiàn)代生物技術(shù)的育種方法成了麻類作物的主流育種技術(shù)。較常見(jiàn)的3種基于生物技術(shù)的育種方法有,一是分子標(biāo)記輔助育種,目前常用的分子標(biāo)記有簡(jiǎn)單重復(fù)序列和單核苷酸多態(tài)性等。這些分子標(biāo)記在麻類作物中主要應(yīng)用于物種演化及親緣關(guān)系研究、群體多樣性研究、標(biāo)記輔助選擇和遺傳圖譜構(gòu)建等。
另一種是利用全基因組選擇(genomic selection,GS),即利用所有可用的標(biāo)記進(jìn)行育種值的預(yù)測(cè)。在GS中,所有的位點(diǎn)、單倍型和標(biāo)記效應(yīng)都是在整個(gè)基因組水平上進(jìn)行估計(jì)的,根據(jù)已知的表型和基因型數(shù)據(jù),以計(jì)算代表性群體中每個(gè)個(gè)體的基因組估計(jì)育種值(genome estimation breeding values,GEBVs)。連鎖不平衡(linkage disequilibrium,LD)在GS方法使用中是一個(gè)非常重要的參數(shù),對(duì)于LD衰減速率快的物種,利用沒(méi)有親緣關(guān)系的自然群體,可以使用數(shù)量較多的標(biāo)記可以在不降低檢測(cè)能力的前提下篩選出用于雜交育種的親本選擇。目前全基因選擇育種已經(jīng)廣泛應(yīng)用于模式作物育種,玉米和水稻等糧食作物育種中有了較深入的研究,但在麻類作物上還鮮有報(bào)道。全基因組選擇育種研究在麻類作物遺傳改良中正處于起步階段,隨著基因型分析成本的降低和統(tǒng)計(jì)方法的發(fā)展,基因組選擇將會(huì)逐步完善,加快我國(guó)麻類作物育種發(fā)展的步伐。
還有一種是利用基因編輯育種。隨著科學(xué)技術(shù)的進(jìn)步,被稱為上帝之手的基因編輯應(yīng)用于遺傳改良,已成為當(dāng)今的作物遺傳改良研究熱點(diǎn)。序列特異性核酸酶,包括鋅指核酸酶(zinc-finger nuclease, ZFN)和轉(zhuǎn)錄激活物樣效應(yīng)核酸酶(transcription activator-like effector nuclease, TALEN)已被初步應(yīng)用于真核生物中進(jìn)行靶向基因組編輯。另一種基于雙鏈斷裂的基因組編輯技術(shù) CRISPR/Cas9系統(tǒng),是在細(xì)菌和古菌群中利用規(guī)則間隔短回文重復(fù)序列(clustered regularly interspaced short palindromic repeats, CRISPR)自適應(yīng)免疫系統(tǒng)的基礎(chǔ)上發(fā)展起來(lái)的,它與ZFN和TALEN相比,操作更簡(jiǎn)單、精確度和效率也都更高。這種新技術(shù)目前已在麻類作物上有所應(yīng)用,如構(gòu)建針對(duì) FAD2 基因的敲除載體,為進(jìn)一步實(shí)現(xiàn)在亞麻植株體內(nèi)對(duì)基因高效率的定點(diǎn)編輯,以及 CRISPR/Cas9系統(tǒng)在亞麻基因組編輯中的應(yīng)用提供參考。
3.3 下一代測(cè)序(NGS)技術(shù)對(duì)麻類作物遺傳改良帶來(lái)新的策略提供可能
在下一代測(cè)序(next generation sequencing, NGS)技術(shù)之前,一個(gè)標(biāo)準(zhǔn)的遺傳連鎖圖譜是基于幾百個(gè)分子標(biāo)記來(lái)構(gòu)建。NGS技術(shù)的發(fā)展與許多其他新技術(shù)相結(jié)合,使得DNA測(cè)序具有高通量和成本低的特點(diǎn),麻類作物基因組測(cè)序完成為基于自然變異的正向遺傳學(xué)策略提供可能。目前SNP標(biāo)記在主要麻類作物遺傳育種研究中的應(yīng)用鮮有報(bào)道,首個(gè)黃麻SNP連鎖圖譜用于對(duì)RIL群體是否抗莖腐病進(jìn)行基因分型,準(zhǔn)確率高達(dá) 91%。這表明,NGS技術(shù)對(duì)麻類作物高密度遺傳圖譜構(gòu)建和重要性狀定位以及標(biāo)記輔助育種帶來(lái)新的策略。
隨著NGS技術(shù)的發(fā)展,我們可以對(duì)麻類作物的不同品種進(jìn)行深度重測(cè)序和全基因組從頭測(cè)序并進(jìn)行等位基因變異分析,通過(guò)序列比對(duì)找到了大量與重要性狀相關(guān)的SNP位點(diǎn),為品質(zhì)改良和新品種選育帶來(lái)了新的科研方法,加快了新品種的育種進(jìn)程。在沒(méi)有參考基因組之前,基于酶切的簡(jiǎn)化基因組測(cè)序(restriction-site associated DNA sequencing, RAD-seq)和基因分型測(cè)序(genotyping by sequencing, GBS)被應(yīng)用于植物中,簡(jiǎn)化基因組測(cè)序技術(shù)具有不依賴于基因組序列,而進(jìn)行高通量的SNP標(biāo)記開(kāi)發(fā)的優(yōu)點(diǎn)。為了解亞麻銹病致病機(jī)制,基于RAD-seq技術(shù)檢測(cè)到的SNP構(gòu)建高密度遺傳圖譜,檢測(cè)到2個(gè)新的致病基因,提高了對(duì)該病原體致病機(jī)制的認(rèn)識(shí);亞麻和苧麻也都利用GBS技術(shù)構(gòu)建了高密度遺傳圖譜,分別定位了株高和纖維相關(guān)QTL。
隨著麻類作物參考基因組圖譜繪制,通過(guò)利用基因組所有染色體上的標(biāo)記和性狀之間的連鎖不平衡(LD),進(jìn)行全基因組關(guān)聯(lián)分析(genome-wide association study, GWAS),能對(duì)自然群體或人工創(chuàng)造群體中的雜表型進(jìn)行基因鑒定。112份苧麻核心種質(zhì)GWAS發(fā)現(xiàn)6個(gè)與分枝數(shù)相關(guān)的候選基因,為研究苧麻候選數(shù)量性狀位點(diǎn)和控制苧麻分枝數(shù)的基因提供信息。亞麻GWAS報(bào)道較多,鑒定出與纖維、亞麻疫病和亞麻種子油含量相關(guān)候選基因,大量豐富了亞麻育種遺傳資源。這些研究反映出,下一代測(cè)序(NGS)技術(shù)對(duì)麻類作物自然群體的優(yōu)良等位基因變異發(fā)掘以及基因組選擇育種帶來(lái)新的策略。
3.4 麻類作物品種改良的發(fā)展趨勢(shì)
目前的麻類作物育種計(jì)劃仍是依賴于將分子標(biāo)記選擇整合到常規(guī)育種方案中的表型選擇,即常規(guī)育種與生物技術(shù)相結(jié)合。過(guò)去,麻類作物的育種目標(biāo)大多是高產(chǎn)優(yōu)質(zhì)的纖維品種培育。但隨著社會(huì)發(fā)展,市場(chǎng)需求與產(chǎn)品多樣化對(duì)麻類育種工作者提出新的挑戰(zhàn),需要選育出高產(chǎn)高效、抗逆抗病、適宜輕簡(jiǎn)化機(jī)械化、優(yōu)質(zhì)專用的多用途麻類作物新品種。
一是麻類纖維用途不能只是局限于紡織業(yè)。由于麻類纖維可自然降解,因此以麻纖維為主要原料可制作環(huán)保型的包裝帶或可降解地膜等。麻類纖維具有纖維強(qiáng)力好的特點(diǎn),其地膜的抗拉強(qiáng)力大于普通地膜,適宜機(jī)械鋪膜,不僅能減少塑料薄膜導(dǎo)致的白色污染,而且能減輕勞動(dòng)強(qiáng)度和生產(chǎn)成本。麻纖維還能用于建筑或家飾板材等用途。近年來(lái)關(guān)于麻纖維復(fù)合材料的研究越來(lái)越多,這是由于與玻璃纖維復(fù)合材料相比麻纖維復(fù)合材料具有密度低、隔音效果好、價(jià)格低和人體親和性好等優(yōu)點(diǎn)。因此培育出更適合用于制作復(fù)合材料的新品種也是當(dāng)下麻類作物育種的一大趨勢(shì)。
二是為了適應(yīng)現(xiàn)代機(jī)械化,麻類作物育種需要滿足輕簡(jiǎn)栽培需要。直播和機(jī)械化等輕簡(jiǎn)栽培技術(shù)能大大降低勞動(dòng)成本,目前已經(jīng)成為水稻、玉米等作物的主流生產(chǎn)技術(shù),然而麻類作物想要適應(yīng)輕簡(jiǎn)栽培還需要克服易倒伏和麻稈易斷等問(wèn)題。為滿足麻類作物適應(yīng)現(xiàn)代機(jī)械化,育種家們需要培育出莖稈粗壯、耐密植、抗倒伏能力強(qiáng)和高產(chǎn)優(yōu)質(zhì)抗病的麻類作物新品種。對(duì)于短日照喜溫類型麻類作物,宜采取“南種北植”來(lái)延長(zhǎng)其營(yíng)養(yǎng)生長(zhǎng)期,提高纖維產(chǎn)量。
三是為了“不與糧爭(zhēng)好地”,培育適合逆境農(nóng)業(yè)的品種。麻類作物在貧瘠土壤的種植,能具有更強(qiáng)的生產(chǎn)潛能,這是棉花所不能比擬的,培育適合逆境農(nóng)業(yè)的新品種不僅僅是要求麻類作物能在不良環(huán)境下保持正常生長(zhǎng),更要穩(wěn)定的達(dá)到高產(chǎn)和優(yōu)產(chǎn)的特點(diǎn)。因此,麻類作物育種工作要更注重耐鹽堿、抗旱、重金屬吸附和抗病蟲(chóng)等性能。
四是根據(jù)不同麻類作物特異性狀,培育不同用途的專用品種。如菜用黃麻、高大麻二酚(CBD)大麻品種、籽用大麻和籽用亞麻等。福建農(nóng)林大學(xué)針對(duì)福建春夏蔬菜淡季的缺口,以中晚熟、高產(chǎn)、食用品質(zhì)優(yōu)、抗炭疽病為選育目標(biāo), 著力開(kāi)展菜用黃麻新品種的選育,最終選育出福農(nóng)系列菜用黃麻,主食嫩莖和幼葉,口感極佳。大麻素作為大麻獨(dú)有的一類次生代謝物,具有重要的醫(yī)用價(jià)值,培育高CBD大麻品種將一直是大麻遺傳改良的重點(diǎn)方向,這也是近年來(lái)大麻產(chǎn)業(yè)蓬勃發(fā)展的重要原因之一。亞麻籽油營(yíng)養(yǎng)豐富,不僅含α-亞麻酸這一人體所必須的脂肪酸,而且亞油酸等不飽和脂肪酸含量高,對(duì)皮膚具有優(yōu)良的親和力和滲透力,能用于護(hù)膚產(chǎn)品原料,為滿足市場(chǎng)需求,培育專用的籽用亞麻是亞麻育種工作必不可少的。
4 展望
從優(yōu)良品種到地方品種和野生近緣種的種質(zhì)資源,將仍然是任何育種計(jì)劃的基礎(chǔ)。隨著麻類作物參考基因組測(cè)序完成,如何高效挖掘這些麻類種質(zhì)資源的遺傳變異和重要應(yīng)用價(jià)值基因位點(diǎn)信息并應(yīng)用到遺傳改良,值得探索。NGS 革命的第一個(gè)效果是降低每個(gè)測(cè)試數(shù)據(jù)的標(biāo)記成本,在瓊脂糖凝膠或毛細(xì)管測(cè)序儀上運(yùn)行的SSRs 或CAPS標(biāo)記要比在高通量平臺(tái)上運(yùn)行的SNPs 昂貴得多。然后,開(kāi)發(fā)一種全基因組選擇的育種方法,如GS,正在成為可能;并將有助于設(shè)計(jì)新的植物品種,不僅應(yīng)用于少數(shù)特定的農(nóng)藝性狀,而且應(yīng)用于基因組中幾乎所有的基因位點(diǎn);以一種成本合理和相對(duì)快速的方式,有望給下一代育種做出顯著的貢獻(xiàn)。目前基因組選擇育種只是在水稻和玉米等主要農(nóng)作物中有所報(bào)道,若能將此育種方法應(yīng)用于麻類作物,必將加快麻類作物育種步伐。預(yù)計(jì)在不久的將來(lái),NGS 技術(shù)、生物信息學(xué)和表型自動(dòng)化等方法在種質(zhì)資源的遺傳多樣性中進(jìn)一步應(yīng)用。建立基于優(yōu)異基因資源挖掘與種質(zhì)創(chuàng)新的高通量基因型-表型數(shù)據(jù)庫(kù),創(chuàng)制種質(zhì)資源管理和共享平臺(tái)。創(chuàng)新并集成分子標(biāo)記輔助選擇、GS、轉(zhuǎn)基因等技術(shù),建立高效的快速育種技術(shù)體系,將徹底改變麻類作物育種策略,以實(shí)現(xiàn)更有效的遺傳改良。
麻類作物作為第5大作物群,其遺傳改良越來(lái)越受益于基因組學(xué)的發(fā)展,通過(guò)基因組學(xué)能發(fā)現(xiàn)一批已知位點(diǎn)與功能的基因資源,從而解決育種親本貧乏和選擇效率低的問(wèn)題,同時(shí)還能供轉(zhuǎn)基因育種使用,能大幅度推動(dòng)主要麻類作物遺傳改良。但應(yīng)該指出的是,這些重要基因以及位點(diǎn)的信息絕大數(shù)是基于或依賴于模式植物的同源基因信息。麻類作物纖維相對(duì)于模式植物而言,有其獨(dú)特之處。同樣地,麻類作物響應(yīng)各種非生物脅迫和氮肥等高效利用等方面也有其特異之處。以紅麻耐鹽堿為例,耐鹽紅麻品種在400mmolL1 的NaCl 濃度下可正常生長(zhǎng)。完成麻類作物參考基因組圖譜繪制,采用正向和反向遺傳學(xué)策略挖掘麻類作物的纖維和重要性狀的作用機(jī)制值得關(guān)注,可開(kāi)展麻類作物種質(zhì)資源形成和演化機(jī)制研究,系統(tǒng)解析纖維產(chǎn)量、纖維品質(zhì)、抗病耐逆等農(nóng)藝性狀形成的分子基礎(chǔ)。主要麻類作物特異性狀的相關(guān)基因有所報(bào)道,但功能基因鑒定仍較困難,如工業(yè)大麻性別相關(guān)的基因,這也與功能標(biāo)記的缺乏有關(guān)。同時(shí),在利用這些基因進(jìn)行麻類作物遺傳改良的過(guò)程中還面臨著遺傳轉(zhuǎn)化體系及其基因編輯體系構(gòu)建等挑戰(zhàn),如在CRISPR-Cas9基因編輯系統(tǒng)中,挖掘適用于特定麻類作物的內(nèi)源 U6 啟動(dòng)子并建立相應(yīng)的基因編輯體系,仍需要摸索與優(yōu)化??朔@些挑戰(zhàn)將能顯著提高主要麻類作物遺傳改良效率。
文章來(lái)源:徐益,張力嵐,祁建民,張列梅,張立武.主要麻類作物基因組學(xué)與遺傳改良:現(xiàn)狀與展望[J/OL].作物學(xué)報(bào):1-35[2021-02-17].http://kns.cnki.net/kcms/detail/11.1809.S.20210125.1444.002.html.
更多閱讀
文章排行
最新文章
Copyright by Ramie Research Institute of Hunan Agricultural University
湖南農(nóng)業(yè)大學(xué)農(nóng)學(xué)院/苧麻研究所 版權(quán)所有 2009-2016 湘ICP備10006838號(hào)-1
